Blog
Orbion Team
When Cell-Free Expression Beats E. coli (and When It's a Waste of Money)

Your target is a single-pass membrane protein that's killed three rounds of BL21 cultures. Every induction either gives you nothing or fills the cells with inclusion bodies, and the one construct that grew turned out to be a frameshifted truncation. A colleague tells you to "just do it cell-free." So you price a kit, see the per-reaction cost, and quietly close the tab.
That instinct — that cell-free is a luxury you can't justify — is right about half the time and badly wrong the other half. Cell-free protein synthesis (CFPS) is not a better E. coli. It's a different tool with a different cost curve, and the question isn't whether it's cheaper per milligram (it usually isn't) but whether your specific bottleneck is one CFPS actually removes.
Key Takeaways
Cell-free wins on toxicity and open chemistry, not cost. With no membrane to poison and no viability to maintain, CFPS expresses proteins that kill the host — toxins, lytic enzymes, proton-coupled transporters — and lets you add nanodiscs, detergents, chaperones, or non-canonical amino acids directly to the reaction.
It's a screening and triage engine, not a production line. Going from linear DNA to protein in 1–2 hours makes CFPS unbeatable for construct triage and DOE-style condition screens. For grams of material, it's the wrong tool.
Per-milligram cost is the dealbreaker. Reagent-grade CFPS runs roughly an order of magnitude more expensive per mg than a shake flask. If your protein expresses fine in BL21, cell-free is money set on fire.
System choice is its own decision. E. coli extract, wheat germ, insect, CHO/mammalian, and the reconstituted PURE system trade yield against folding competence, disulfides, and glycosylation. Picking the wrong lysate wastes the run.
No native glycosylation, hard scale-up economics. Standard lysates don't build human N-glycans, and the cost curve that makes CFPS great for screening makes it punishing for production. Know your endpoint before you start.

Why Cell-Free Exists: The Constraint It Removes
Every in-cell expression system carries the same baggage: the cell has to stay alive. That single requirement drives most of the failures protein engineers fight. A protein that punches holes in the inner membrane gets selected against. A construct that drains the host's amino acid pool slows growth until revertants take over. A disulfide-rich domain hits the reducing cytoplasm and collapses into inclusion bodies.
Cell-free protein synthesis throws the cell away and keeps only the machinery. You lyse the cells, clear the debris, and add back the transcription-translation apparatus — ribosomes, tRNAs, aminoacyl-tRNA synthetases, energy regeneration, and your DNA template. The reaction is an open tube. Nothing has to grow, divide, or survive [1].
That openness is the whole point. You control the chemistry directly. Reducing or oxidizing redox potential, chaperone cocktails, detergents, lipid nanodiscs, unusual cofactors, isotope-labeled or non-canonical amino acids — all of it goes straight into the reaction at the concentration you choose. In a living cell, none of that is yours to set.
The core tension: this freedom is expensive. The reagents that replace the cell — purified energy mix, the extract itself, the template — cost real money per microliter, and the reaction volume is small. CFPS buys you control over the protein's environment and pays for it in dollars per milligram. The entire decision comes down to whether your problem is one that control solves.

When Cell-Free Genuinely Wins
CFPS earns its cost in a handful of specific situations. If your problem isn't on this list, you probably don't need it.
1. Toxic and lethal proteins
This is the cleanest win. Restriction endonucleases, pore-forming toxins, membrane-disrupting peptides, and lytic enzymes have no host to kill in a cell-free reaction, so they express at full yield instead of selecting for non-expressing escape mutants [2]. CFPS has produced restriction endonucleases, cytolethal distending toxin, and other proteins that are functionally impossible to make in living E. coli without elaborate tight-repression tricks.
Diagnostic question: Does your protein's biological activity attack the host that's supposed to make it? If yes, CFPS removes the conflict entirely.
2. Membrane proteins — with the right additive
Membrane proteins are CFPS's headline application, but with a critical caveat: the cell-free reaction has no membrane. A nascent transmembrane helix emerging into aqueous lysate will aggregate unless you give it somewhere to go. You have to supply that hydrophobic environment yourself.
Three modes work:
Detergent (D-CF): micelles in the reaction capture the protein as it's made. E. coli-based systems have produced structurally intact mammalian membrane proteins — using detergent or supplied membrane fragments — at roughly 0.1–1.0 mg per mL of reaction, enough for downstream structural work [3].
Nanodiscs / preformed bilayers: a lipid bilayer disc gives a near-native environment co-translationally — often the difference between folded and aggregated for a GPCR or transporter.
Precipitate (P-CF) then resolubilize: express without additive, pellet, and solubilize afterward. Cheapest, but you're back to refolding risk.
The win is that you screen detergents and lipids in the synthesis reaction itself, in parallel, in an afternoon — instead of expressing, purifying, and re-screening solubilization conditions over weeks. Eukaryotic membrane proteins with disulfides or complex folds often do better in wheat-germ CFPS, which pairs high translation efficiency with a eukaryotic ribosome [5][6].
Diagnostic question: Is your membrane-protein bottleneck finding the right detergent/lipid? CFPS turns that from a serial slog into a parallel screen. If you already know your detergent and need grams, it doesn't help.
3. Fast construct triage and DOE screening
From linear PCR product to detectable protein in 1–2 hours, with no cloning, no transformation, no overnight cultures [2]. That speed changes what's affordable to test. You can take 48 construct variants — different boundaries, tags, fusion partners, codon schemes — amplify them by PCR, and ask which ones make soluble protein the same day.
This is the highest-value, most underused application. Not as a production method, but as a filter that tells you which constructs deserve a real prep. You burn a small amount of reagent to avoid weeks of cloning dead constructs.
Diagnostic question: Do you have many constructs and no idea which will express? CFPS ranks them in a day.
4. Non-canonical amino acid incorporation
The open reaction is ideal for site-specific ncAA incorporation. You add the orthogonal aminoacyl-tRNA synthetase, the suppressor tRNA, and the unnatural amino acid at controlled concentrations, free of the competing endogenous machinery and uptake limits that constrain in-cell systems [4]. The reconstituted PURE system goes further: because you build it from defined components, you can omit release factor 1 to make stop-codon suppression dramatically more efficient [7]. For one or two precisely placed click handles, fluorophores, or photo-crosslinkers, CFPS is the most direct route.
5. Linear-template and library work
Because no transformation is needed, CFPS expresses straight from PCR products. That makes it natural for expressing computationally designed libraries, scanning point variants, or any workflow where cloning every member is the bottleneck rather than the protein itself.
When Cell-Free Is a Waste of Money
Reach for CFPS in the wrong situation and you've paid a premium for nothing.
Your protein already expresses in E. coli
If BL21 gives you 20 mg/L of soluble, folded protein, stop. A shake flask is one of the cheapest unit operations in molecular biology. CFPS reagents cost roughly an order of magnitude more per milligram of product. There's no prize for making an easy protein the hard way.
You need grams, not milligrams
CFPS reactions are small and the reagents are the cost driver, so the economics run backwards from fermentation. E. coli extract reactions can be scaled up to deliver milligram quantities of purified membrane protein per run [3], but a preparative cell-free reaction is an expensive thing to run, and it doesn't get cheaper per mg as you scale the way a bioreactor does. For multi-gram material, in-cell fermentation wins on cost every time.
You need authentic human glycosylation
Standard CFPS lysates don't build complex human N-glycans. E. coli extract has no glycosylation machinery at all; even eukaryotic lysates lack the intact, compartmentalized Golgi pathway that produces native glycan structures. For a therapeutic antibody or any glycoprotein whose function or developability depends on its glycan, a CHO or HEK cell line remains the answer. CHO-lysate cell-free systems exist and handle disulfides and some difficult-to-express targets [8], but they are not a substitute for in-cell glycan processing.
You haven't matched the system to the protein
Treating "cell-free" as one thing is a common, expensive mistake. An E. coli extract won't fold a disulfide-rich eukaryotic protein well in its default reducing state; a wheat-germ system gives you eukaryotic folding but at different yield and cost. Pick the wrong lysate and the run fails for reasons that have nothing to do with CFPS being right for the job.

Choosing a Lysate System
The five common systems are not interchangeable. Match the system to what your protein needs to fold.
System | Ribosome | Best at | Disulfides | Glycosylation | Relative yield | Relative cost |
|---|---|---|---|---|---|---|
E. coli extract | Prokaryotic | Toxic proteins, bacterial targets, speed, scale-up | No (needs oxidizing/PDI add-back) | None | High | Lowest |
Wheat germ | Eukaryotic | Complex eukaryotic & membrane proteins, multidomain | Yes | Limited | High (eukaryotic) | Medium |
Insect (Sf21) | Eukaryotic | Eukaryotic proteins, some PTMs, microsomes | Yes | Partial | Medium | Medium-high |
Mammalian / CHO | Eukaryotic | "Difficult-to-express" human proteins, native-like folding | Yes | Partial (no full Golgi) | Lower | High |
PURE (reconstituted) | Prokaryotic | ncAA work, defined background, minimal nucleases | Tunable | None | Low (~160 µg/mL/h) | Highest |
A few honest notes on this table. Wheat germ pairs the highest translation efficiency among eukaryotic cell-free options with a eukaryotic ribosome, which is why it carries a strong track record for membrane proteins and large assemblies [5][6]. The PURE system trades yield for control — built from defined purified components, it gives you a clean, nuclease-poor, RNase-poor background that's ideal for ncAA and minimal-system work, at roughly 160 µg/mL/h in batch [7]. Prokaryotic and eukaryotic systems each have genuine pros and cons; there is no universally best lysate [8].
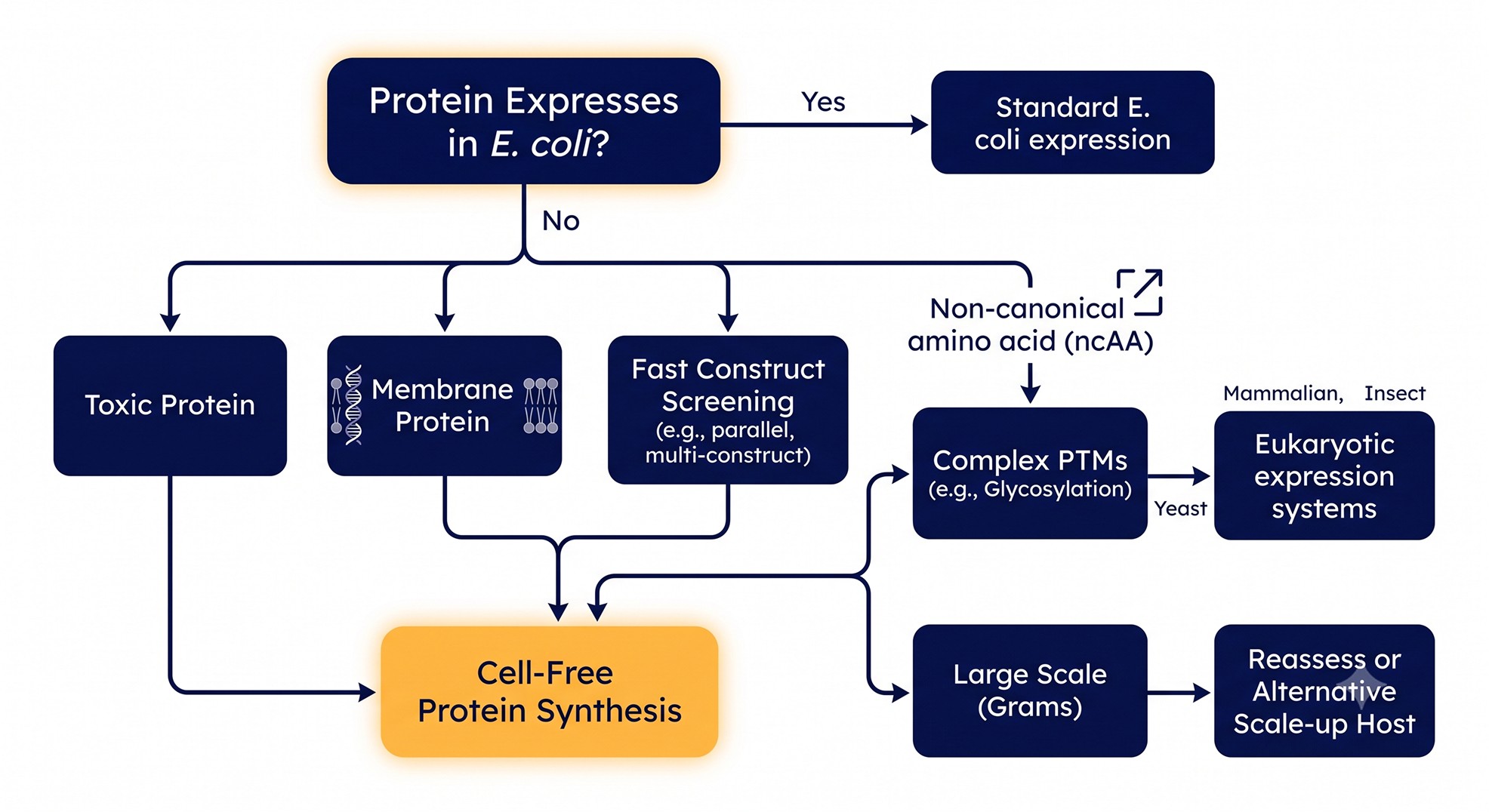
Decision Tree: Cell-Free or Not

Case Study: Triage Before a GPCR Crystallography Campaign
Problem: A structural team had a Class A GPCR target that refused to express in E. coli at usable levels. Inclusion bodies dominated, and the one detergent-solubilized prep that purified was monodisperse one week and aggregated the next. They were about to commit a baculovirus campaign to a single construct chosen essentially by guesswork — roughly six weeks and substantial reagent cost per construct tested.
Analysis: The real bottleneck wasn't the host. It was that nobody knew which of eight construct designs (varying N-terminal truncation, fusion partner, and detergent compatibility) would fold, and the in-cell test loop was too slow to find out. Each guess cost weeks.
Solution: They ran all eight constructs in a wheat-germ cell-free reaction directly from PCR products, screening three detergents and a nanodisc condition per construct in parallel — 32 micro-reactions in a single afternoon. Solubility and monodispersity were read out the same day.
Outcome: Two of eight constructs gave soluble, monodisperse protein; one was clearly best with a specific detergent. The team carried that construct into baculovirus instead of gambling. They spent a day and a small amount of reagent to avoid an estimated multi-week dead end, and the chosen construct expressed and purified on the first in-cell attempt. CFPS never made a milligram of crystallization-grade protein here — it just told them which prep was worth running.
Practical Checklist Before You Run CFPS
Before committing reagents, verify:
You've confirmed E. coli actually fails — not just that it's inconvenient. CFPS only pays off when in-cell expression is genuinely blocked.
You know your endpoint: screening/triage (CFPS shines) vs. grams of production (it doesn't).
You've matched the lysate to the fold: prokaryotic vs. eukaryotic ribosome, disulfide handling, PTM needs.
For membrane proteins, you've chosen a folding additive — detergent, nanodisc, or preformed bilayer. A bare reaction will aggregate them.
For disulfide-bonded targets, you've set the redox environment (oxidizing buffer, DsbC/PDI add-back) rather than using a default reducing mix.
You don't need authentic human glycosylation — or you've accepted that a cell line is the real answer.
You've priced it per milligram of final product, not per reaction, against a shake flask.
The Economics
Numbers below are order-of-magnitude planning figures; they vary widely with system, scale, and whether you make extract in-house or buy kits. Use them to reason about which side of the line your project sits on, not as a quote.
Approach | Time to protein | Typical yield | Relative cost per mg | Best for |
|---|---|---|---|---|
E. coli shake flask | 1–3 days | 1–50+ mg/L culture | Lowest | Anything that expresses |
E. coli extract CFPS | 1–2 hours | ~0.1–1.0 mg/mL reaction | ~10× a shake flask | Toxic proteins, fast triage, scalable membrane work |
Wheat-germ CFPS | Hours | High (eukaryotic) | Medium-high | Complex/eukaryotic & membrane proteins |
CHO/mammalian CFPS | Hours | Lower | High | Difficult human proteins, native-like folding |
PURE system | 1–2 hours | ~160 µg/mL/h | Highest | ncAA, defined-background work |
Baculovirus / CHO cell line | Weeks | High at scale | Low at scale, high per construct tested | Glycoproteins, gram-scale production |
ROI consideration: CFPS rarely wins on cost per milligram. It wins on cost per decision. A day of cell-free screening that kills six dead-end constructs before a six-week baculovirus campaign pays for itself many times over — even though every one of those milligrams was "expensive." Frame the spend against the weeks of in-cell work it lets you skip, not against the price of a shake flask.
Bottom Line
Cell-free expression is a control-and-speed instrument, not a cheap one. Use it when your protein is toxic, when a membrane target needs co-translational detergent or nanodiscs, when you need to triage many constructs fast, or when you're placing non-canonical amino acids — and don't use it when E. coli already works, when you need grams, or when you need authentic human glycans. Then pick the lysate that matches your protein's folding requirements, because "cell-free" is five different systems wearing one name.
How Orbion Helps
The hard part of this decision usually isn't running the reaction — it's predicting, before you spend anything, whether your target is toxic-membrane-and-fussy or boringly easy. That triage is exactly where Orbion fits.
Run a sequence through Characterization and AstraSUIT returns experimental-suitability and host-association predictions — including membrane type and subcellular location — so you can see upfront whether a target leans toward the bacterial-host or eukaryotic-host route, and whether membrane topology is going to be the problem. AstraUNFOLD maps transmembrane topology, disorder, and amyloid propensity, the three signals that most often decide whether a cell-free reaction needs nanodiscs or fails outright. The Bench module's Rate of Ease score — derived from solubility, disorder, aggregation, and membrane-topology predictions — gives you a single triage number for which targets actually justify the cell-free premium versus which will express in a flask.
When you've decided cell-free is warranted, the Bench module's Custom Design lets you set the expression system explicitly — E. coli, insect, mammalian, cell-free, or auto-recommend — and generates construct-aware protocols built around that choice, with the assembled protein and codon-optimized DNA sequence ready for a linear-template reaction. Pair that with the Design tab to generate and score construct variants before you commit reagents, so the constructs you put into a cell-free triage screen are the ones already most likely to fold.
Ready to find out whether your stubborn target is worth the cell-free premium? Run it through Orbion's Characterization module and read the Rate of Ease before you order a single kit.
References
Dondapati SK, Stech M, Zemella A, Kubick S. (2020). Cell-Free Protein Synthesis: A Promising Option for Future Drug Development. BioDrugs, 34(3):327-348. Link
Jin X, Hong SH. (2018). Cell-free protein synthesis for producing 'difficult-to-express' proteins. Biochemical Engineering Journal, 138:156-164. Link
Shinoda T, Shinya N, Ito K, et al. (2016). Cell-free methods to produce structurally intact mammalian membrane proteins. Scientific Reports, 6:30442. Link
Cui Z, Johnston WA, Alexandrov K. (2020). Cell-Free Approach for Non-canonical Amino Acids Incorporation Into Polypeptides. Frontiers in Bioengineering and Biotechnology, 8:1031. Link
Harbers M. (2014). Wheat germ systems for cell-free protein expression. FEBS Letters, 588(17):2762-2773. Link
Fogeron ML, Lecoq L, Cole L, Harbers M, Böckmann A. (2021). Easy Synthesis of Complex Biomolecular Assemblies: Wheat Germ Cell-Free Protein Expression in Structural Biology. Frontiers in Molecular Biosciences, 8:639587. Link
Shimizu Y, Inoue A, Tomari Y, et al. (2001). Cell-free translation reconstituted with purified components. Nature Biotechnology, 19(8):751-755. Link
Zemella A, Thoring L, Hoffmeister C, Kubick S. (2015). Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem, 16(17):2420-2431. Link
Book a 20-Minute Demo
Sign up free for unlimited Overview runs — summary, sequence-based analysis, homology search. For the full Characterization — PTMs, binding sites, stability variants, construct design — book a demo and we'll run your target live.
