Blog
Orbion Team
Why Does My Protein Elute in the Void Volume on SEC
You load your "pure" protein onto a Superdex 200 column, expecting a nice peak at 70 kDa. Instead, the entire sample elutes in the void volume—the dead space before the column even starts separating. Your protein is either enormous, aggregated, or both. And the purification you thought was done isn't.
The void volume peak is one of the most common and most misinterpreted results in protein biochemistry. Here's what it means and what to do about it.
Key Takeaways
Void volume = your protein is too large for the column's separation range, either because it's a genuine large complex, an aggregate, or is interacting with the column matrix
Aggregation is the most common cause—not that your protein is secretly a megadalton complex
Intrinsically disordered regions inflate apparent size by 1.5–3x, pushing even monomeric proteins into the void
Column interactions (not just size) can cause void elution: charge repulsion from the matrix pushes proteins through faster
The fix depends on the cause: aggregation needs construct/buffer redesign; IDR inflation needs SEC-MALS for true MW; column interactions need salt adjustment
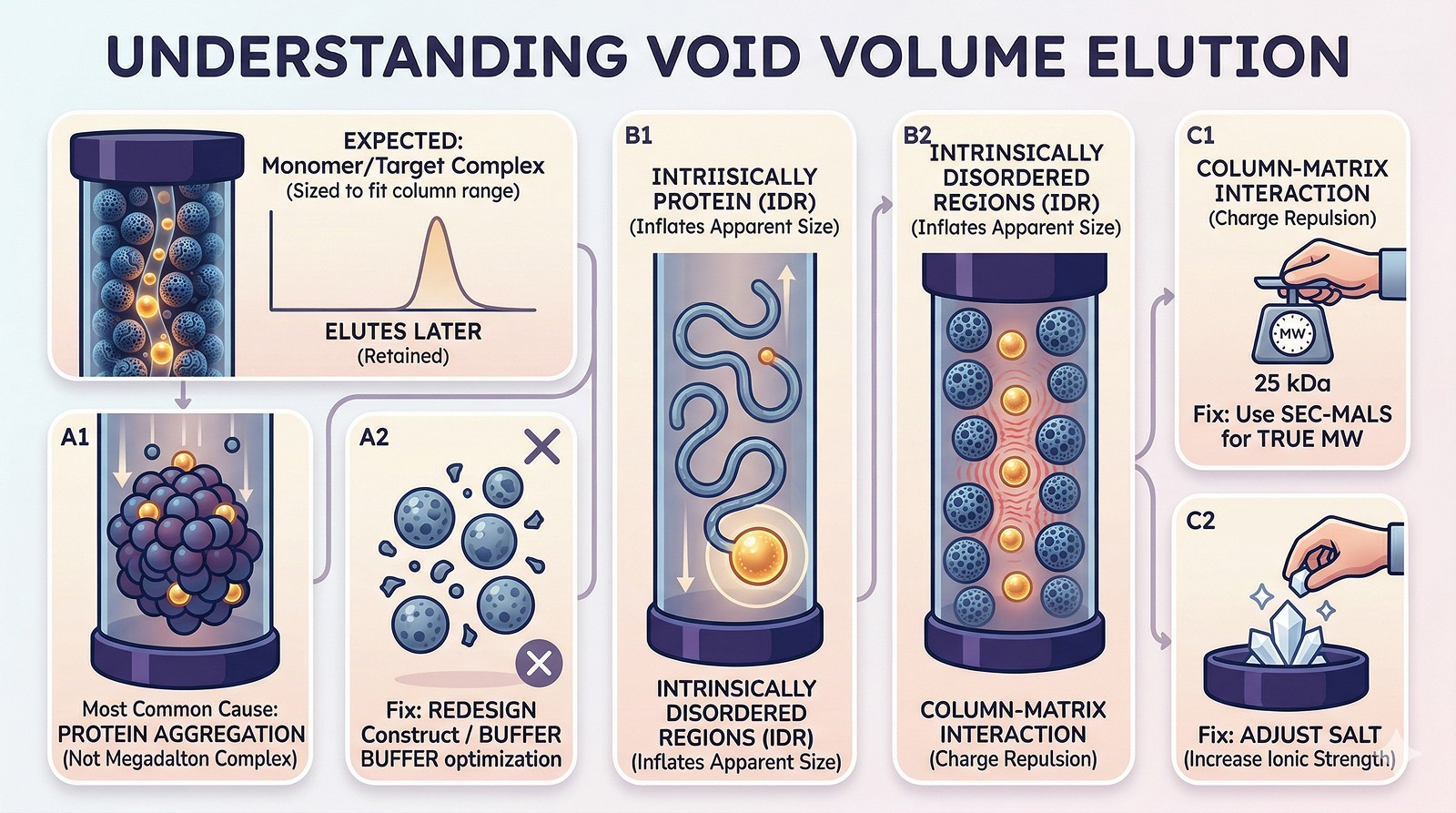
What the Void Volume Actually Is
The void volume (V₀) is the volume of buffer between the resin beads—the space that molecules too large to enter any pores pass through without separation.
Column | Separation Range | Void Volume |
|---|---|---|
Superdex 75 | 3–70 kDa | Anything >70 kDa elutes in void |
Superdex 200 | 10–600 kDa | Anything >600 kDa elutes in void |
Superose 6 | 5–5,000 kDa | Anything >5,000 kDa elutes in void |
First check: Is your protein simply larger than the column's range? A 200 kDa complex on a Superdex 75 will always elute in the void. That's not a problem—it's the wrong column.
The Five Causes of Void Volume Elution
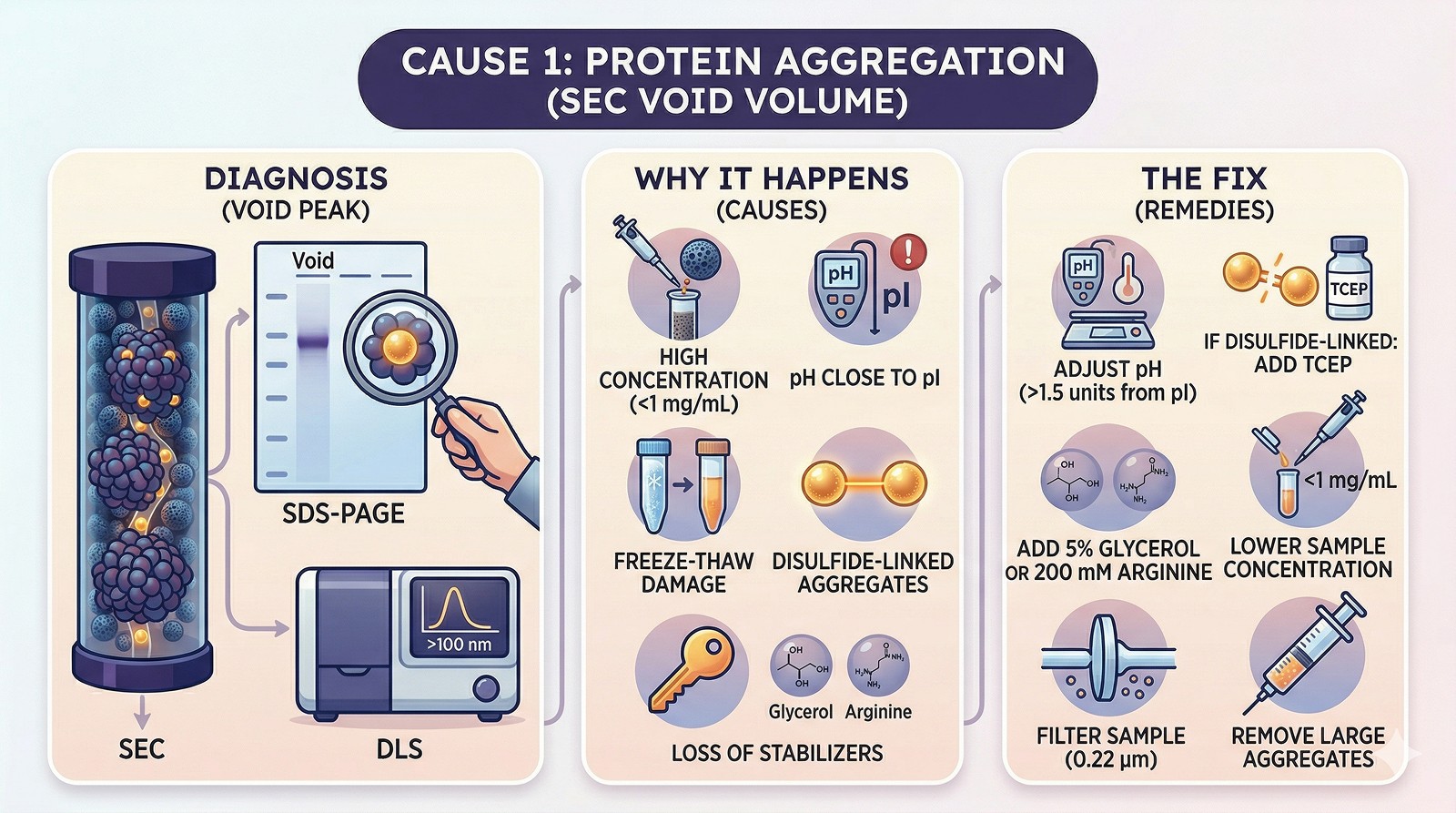
Cause 1: Aggregation (Most Common)
Diagnosis:
Protein was monomeric/oligomeric previously but now elutes in the void
SDS-PAGE of the void peak shows your protein (not contaminants)
Dynamic light scattering shows particles >100 nm
Why it happens:
Concentration-dependent aggregation during loading
Buffer incompatibility (pH near pI, low ionic strength)
Freeze-thaw damage
Oxidation of surface cysteines → disulfide-linked aggregates
Removal of stabilizing additives (glycerol, arginine, detergent)
Fix:
Run non-reducing SDS-PAGE: if higher MW bands appear, aggregation is disulfide-mediated → add TCEP
Lower the loading concentration (<1 mg/mL)
Add 5% glycerol or 200 mM arginine to the running buffer
Check pH relative to pI (stay >1.5 units away)
Filter sample through 0.22 µm before loading to remove large aggregates
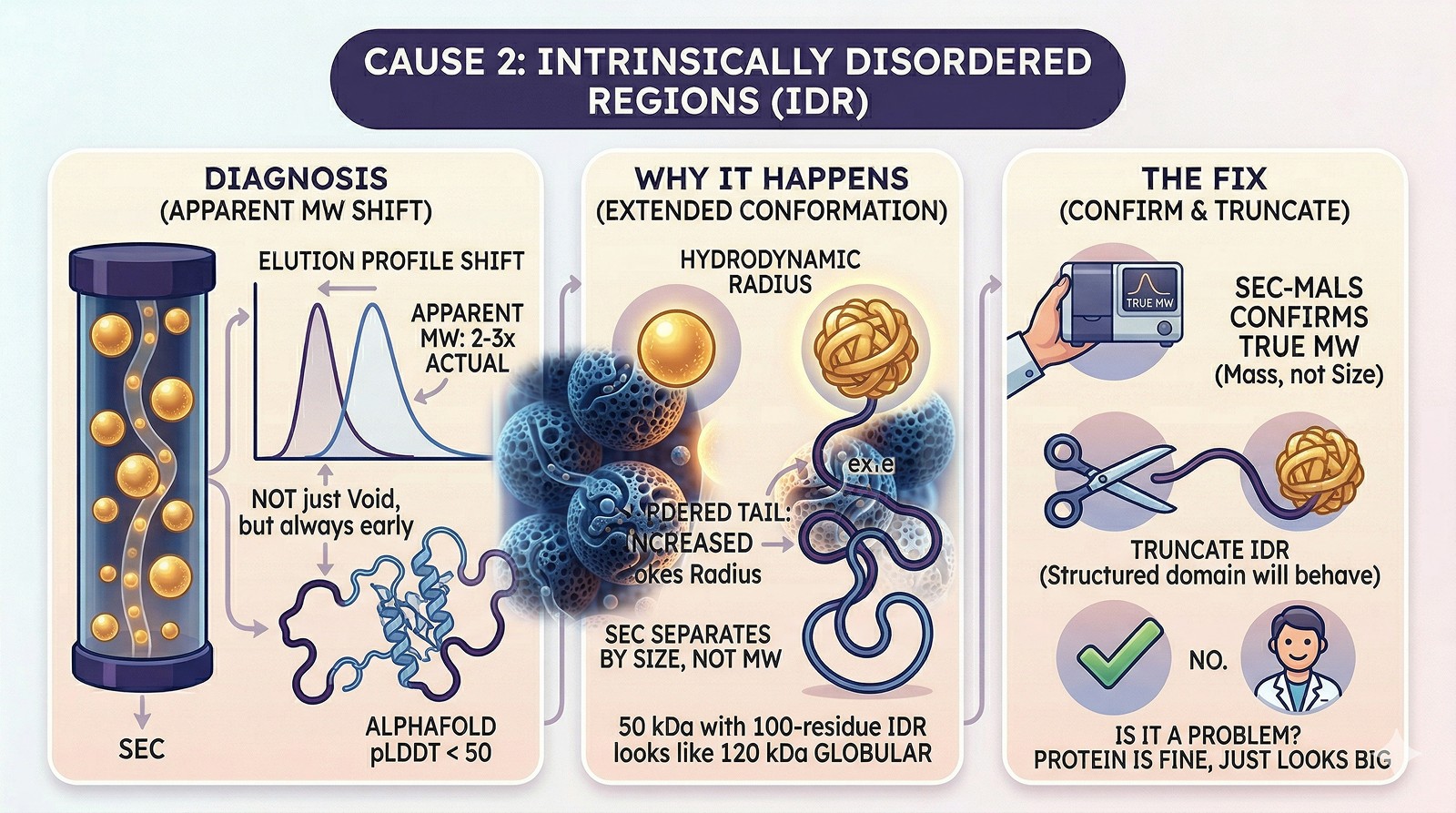
Cause 2: Intrinsically Disordered Regions
Diagnosis:
Protein consistently elutes earlier than expected MW (not just in void—but apparent MW is 2–3x actual)
AlphaFold model shows long stretches of pLDDT < 50
SEC-MALS gives true MW matching the monomer, despite early elution
Why it happens:
Disordered regions adopt extended conformations, increasing the protein's hydrodynamic radius
SEC separates by size (Stokes radius), not molecular weight
A 50 kDa protein with a 100-residue disordered tail can have the Stokes radius of a 120 kDa globular protein
Fix:
Use SEC-MALS to confirm true molecular weight (it measures mass, not size)
If the IDR isn't needed, truncate it—the structured domain will behave normally on SEC
This is NOT a problem if you know it's happening—the protein is fine, it just looks big
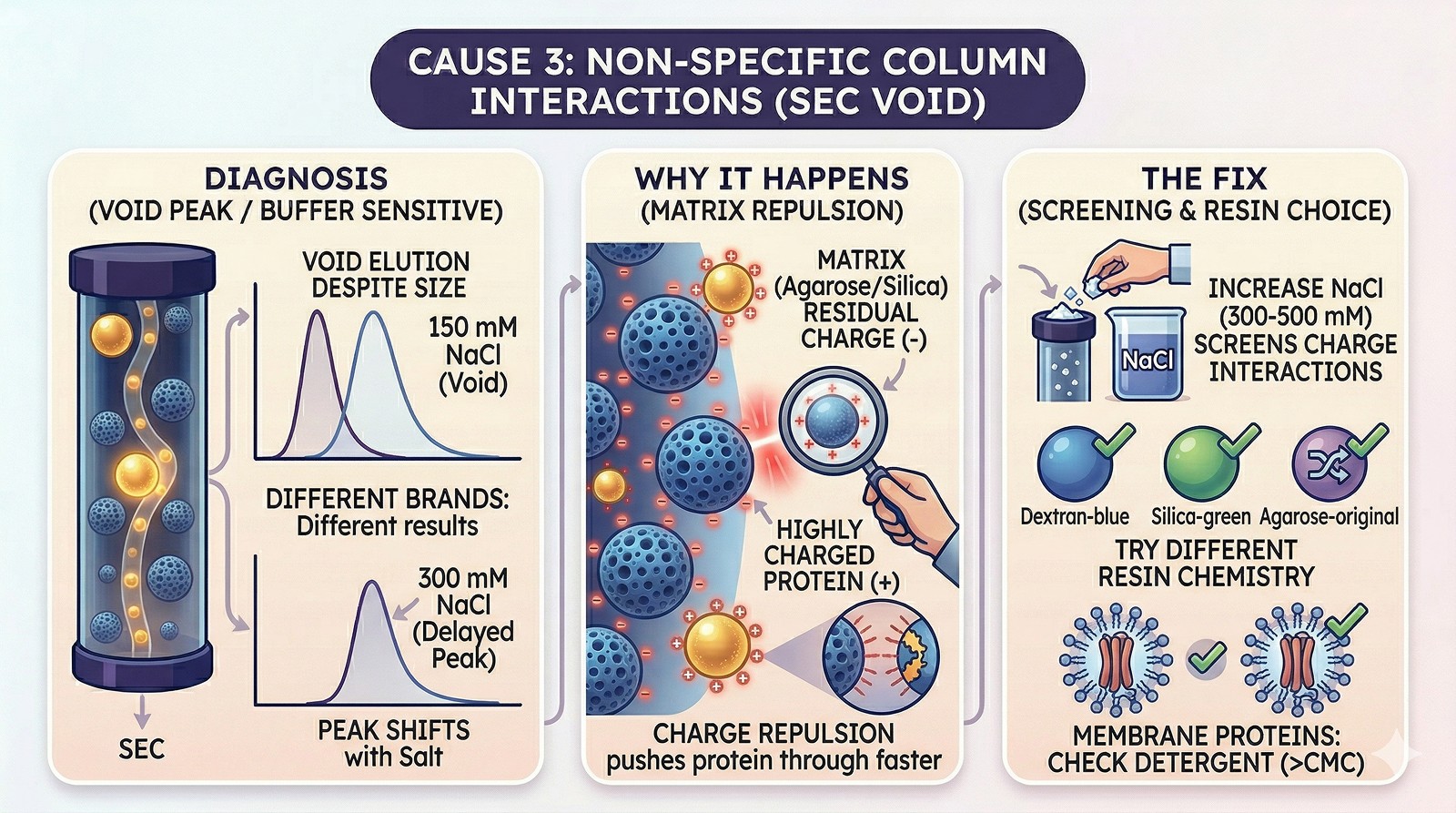
Cause 3: Non-Specific Column Interactions
Diagnosis:
Protein elutes in or near the void even though it's well within the separation range
Changing running buffer salt concentration changes elution position
Protein elutes normally on a different brand/type of column
Why it happens:
Some SEC resins have residual charges (slightly negative for agarose-based resins)
Highly positively charged proteins are repelled from the matrix → elute earlier
Highly negatively charged proteins may be attracted → elute later (or stick)
Detergent-solubilized membrane proteins interact with the matrix hydrophobically
Fix:
Increase NaCl to 300–500 mM in the running buffer (screens charge interactions)
Try a different resin chemistry (dextran vs agarose vs silica)
For membrane proteins, ensure sufficient detergent in the buffer (above CMC)
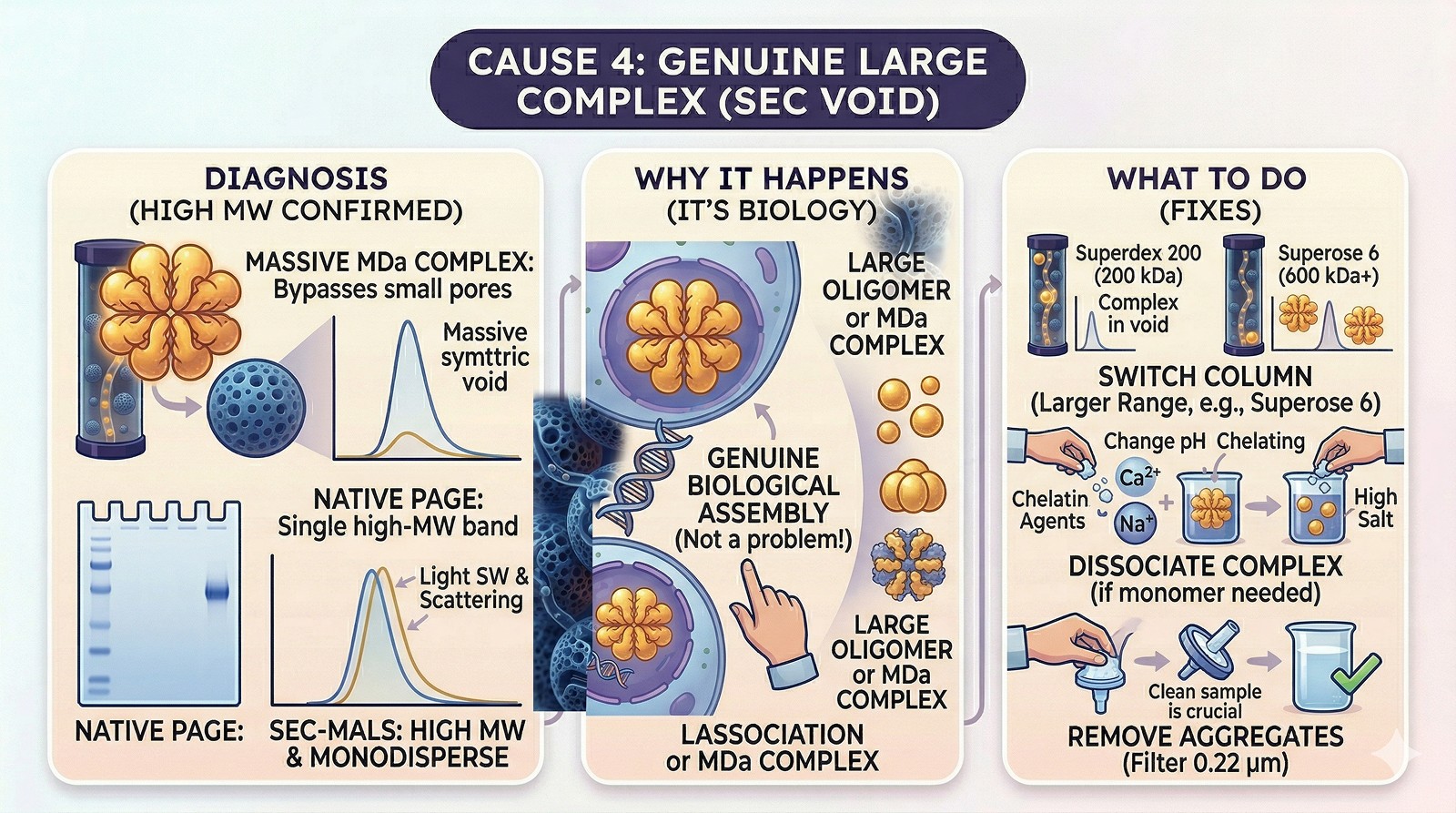
Cause 4: Genuine Large Complex
Diagnosis:
Protein is known to form a large oligomer or part of a megadalton complex
SEC-MALS confirms high molecular weight
Native PAGE shows a single, high-MW band
The void peak is symmetrical and monodisperse (DLS shows a single population)
This isn't a problem—it's biology. Your protein genuinely forms a large complex.
What to do:
Switch to a column with a larger separation range (Superose 6 for >600 kDa)
If you need the monomer, find conditions that dissociate the complex (high salt, different pH, chelating agents for metal-dependent assembly)
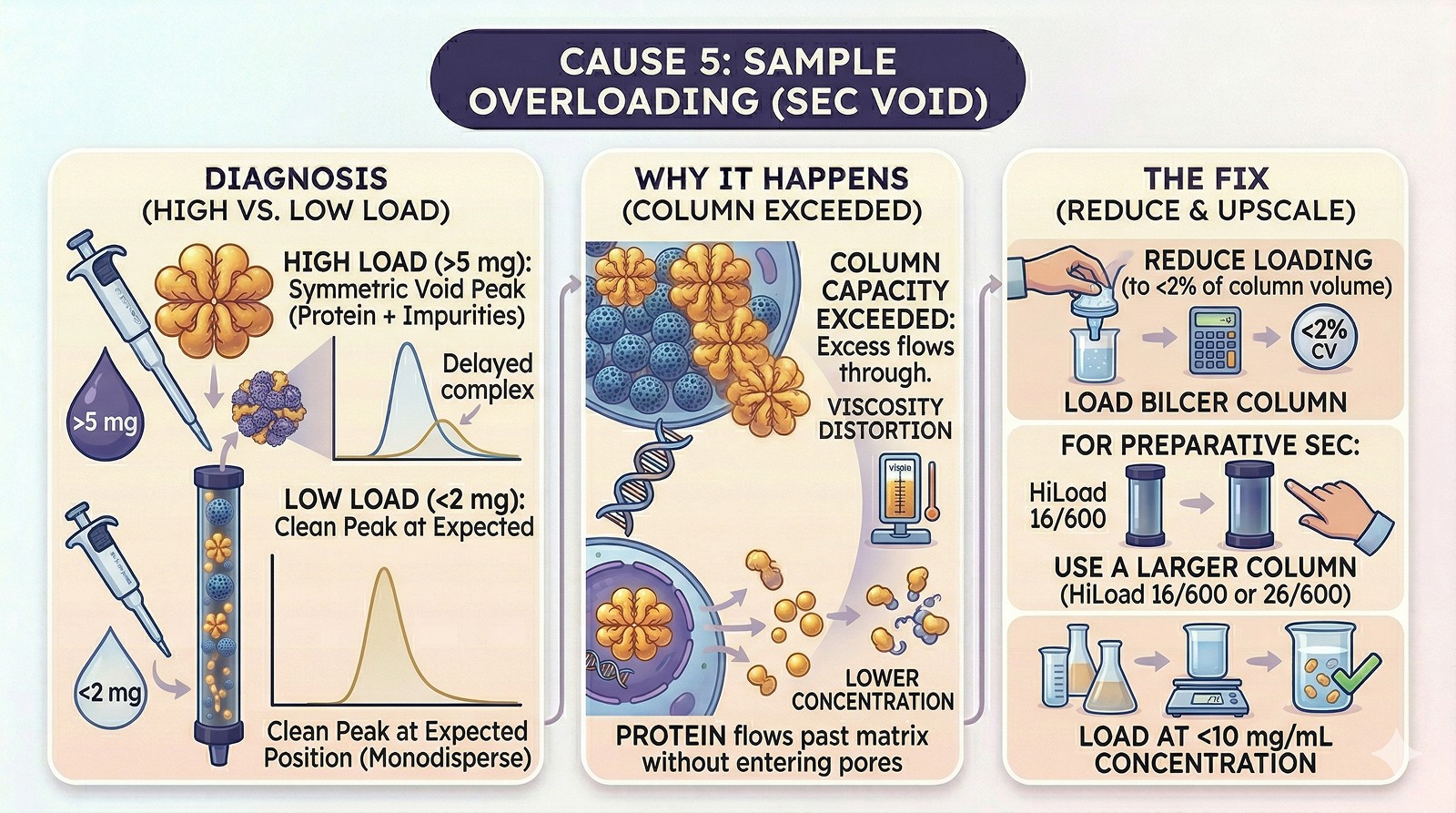
Cause 5: Sample Overloading
Diagnosis:
Void peak appears only when loading >5 mg on a standard analytical column
Lower loads give a clean peak at the expected position
The void peak contains your protein plus impurities
Why it happens:
Column capacity is exceeded—excess protein flows through without entering pores
Viscosity effects at high concentration distort the separation
Fix:
Reduce loading to <2% of column volume
For preparative SEC, use a larger column (HiLoad 16/600 or 26/600)
Load at <10 mg/mL concentration
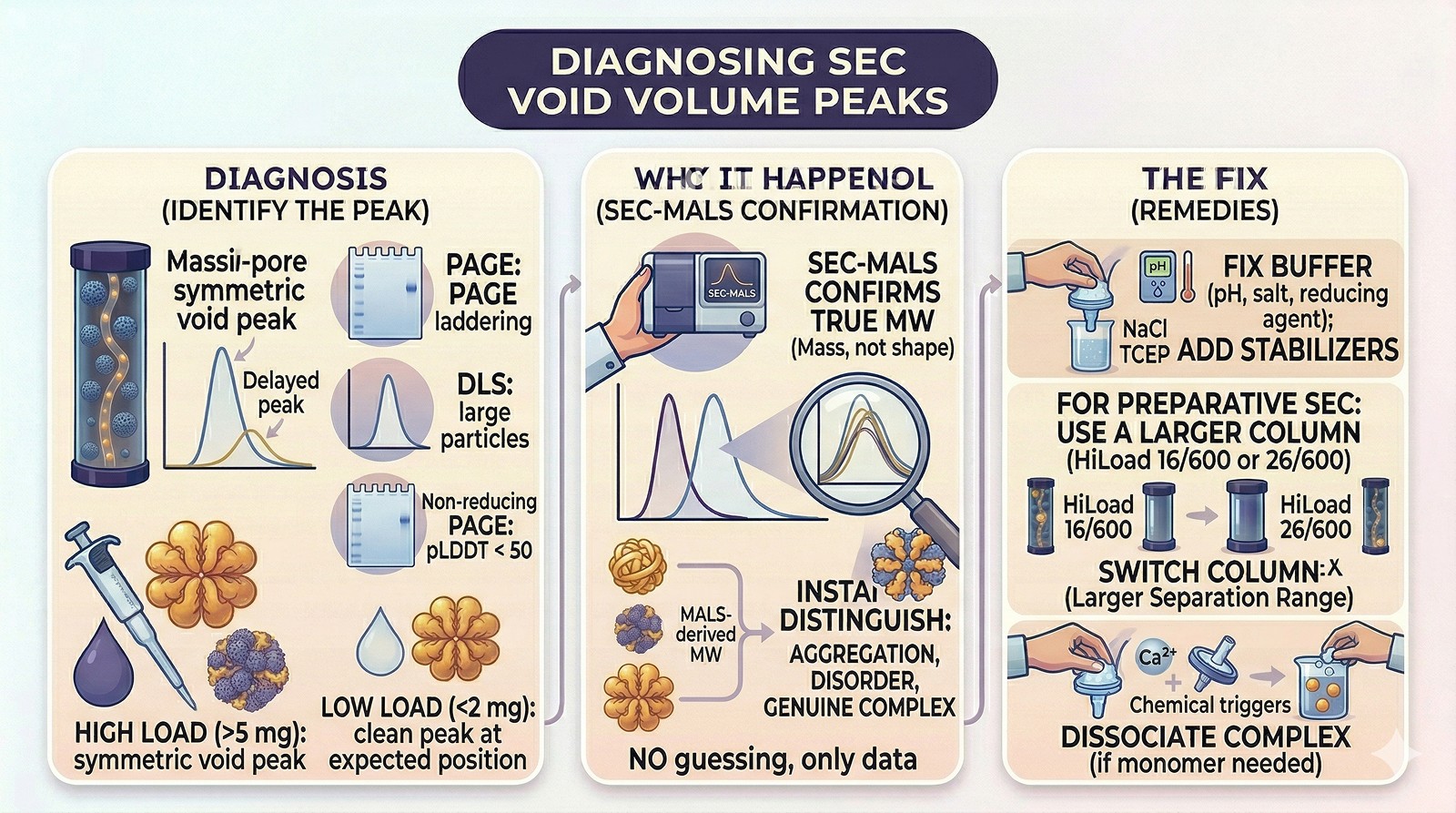
The Diagnostic Flowchart
The Bottom Line
Void Volume Cause | How to Identify | Solution |
|---|---|---|
Aggregation | Non-reducing PAGE shows laddering; DLS shows large particles | Fix buffer (pH, salt, reducing agent); add stabilizers |
Disordered regions | SEC-MALS MW matches monomer; AlphaFold pLDDT < 50 | Truncate IDRs or accept the elution profile |
Column interactions | Elution shifts with salt concentration | Increase NaCl to 300–500 mM |
Genuine large complex | SEC-MALS confirms high MW; monodisperse | Use Superose 6; or dissociate |
Sample overload | Void peak only at high loading | Reduce load amount |
The critical tool: SEC-MALS. If you're seeing void volume peaks and you don't have access to multi-angle light scattering, you're guessing. SEC-MALS tells you the actual molecular weight independent of shape, instantly distinguishing aggregation from disorder from genuine complexes.
Diagnosing Aggregation Before SEC
Orbion flags aggregation risk before you reach the column. AstraUNFOLD predicts per-residue disorder probability and amyloidogenicity—identifying regions that drive aggregation when exposed. The PAE Insight Engine reveals domain boundaries, helping you design constructs that exclude disordered tails inflating your Stokes radius. Combined with Bench module protocols optimized for buffer conditions matched to your protein's properties, you can address the root cause rather than troubleshooting at the SEC stage.
References
Fekete S, et al. (2014). Theory and practice of size exclusion chromatography for the analysis of protein aggregates. Journal of Pharmaceutical and Biomedical Analysis, 101:161-173. Link
Some D, et al. (2019). Characterization of proteins by size-exclusion chromatography coupled to multi-angle light scattering (SEC-MALS). Journal of Visualized Experiments, (148):e59615. Link
Amartely H, et al. (2018). Coupling multi-angle light scattering to ion exchange chromatography (IEX-MALS) for protein characterization. Scientific Reports, 8:6907. Link
Book a 20-Minute Demo
Sign up free for unlimited Overview runs — summary, sequence-based analysis, homology search. For the full Characterization — PTMs, binding sites, stability variants, construct design — book a demo and we'll run your target live.
