You spent three months optimizing expression, two weeks tuning grids, and a 48-hour data collection on the Krios. The 2D classes look gorgeous. But when you run 3D refinement, the map is smeared in one direction—a top view that's crisp from above and stretched into pancake densities from the side. The FSC says 3.2 Å, but you can't trace a side chain. Welcome to preferred orientation: the silent killer of cryo-EM projects.
Preferred orientation is one of the most common reasons high-resolution-on-paper maps cannot be interpreted in practice. It is also one of the most fixable problems—if you correctly diagnose the underlying cause before you reach for a fix.
Key Takeaways
-
Preferred orientation is a sampling problem, not a resolution problem—global FSC can lie spectacularly
-
The air-water interface is the primary culprit in nearly all cases, denaturing or orienting particles via exposed hydrophobic patches
-
Diagnose before fixing: Euler angle distributions, 3D FSC, and sphericity index reveal the underlying geometry
-
Six strategies exist, ranging from "collect at 30° tilt today" to "redesign the construct"—each with distinct trade-offs in time, cost, and risk
-
Surface chemistry is engineerable: hydrophobic patches, dipole moments, and asymmetric Fab handles can all be predicted and modified before you ever freeze a grid
What Preferred Orientation Actually Is
The textbook definition: particles in vitrified ice adopting a non-uniform distribution of Euler angles, such that some orientations are over-represented and others are missing. The physical reality is more uncomfortable.
Single-particle cryo-EM reconstructs a 3D Coulomb-potential map by combining many 2D projections of the same molecule viewed from different angles. The central section theorem says each 2D image fills in one plane of 3D Fourier space passing through the origin. If your particle views are uniformly distributed on the sphere, you fill Fourier space evenly. If they cluster, you fill some directions densely and others not at all.
This is a sampling problem in the strict mathematical sense. The reconstruction algorithm cannot invent information about Fourier components it never measured. It can interpolate, regularize, or apply priors—but the resulting map will be smeared along the under-sampled direction. That smearing manifests in real space as elongated densities, broken side-chain rotamers, and rod-like helices.
The Air-Water Interface Problem
A typical cryo-EM grid hole is ~20-100 nm thick at the center, and ~200-500 nm at the edges. Particles in this thin film of buffer encounter two air-water interfaces—one above, one below. These interfaces are aggressively hydrophobic, and during the ~1 second between blotting and plunging, particles diffuse to and adsorb onto them with surprising efficiency.
Noble et al. (2018) used fluorescence and tomography to estimate that >90% of particles in standard grids contact the air-water interface at least once before vitrification (Noble et al., 2018). D'Imprima et al. (2019) showed that for many proteins this contact is not benign—it denatures particles partially or fully, and even when it does not, it orients them with their hydrophobic faces pinned against the interface (D'Imprima et al., 2019).
Why Particles Pick Favorite Orientations
Three molecular features drive orientation bias:
-
Hydrophobic surface patches. Membrane-protein cytoplasmic faces, exposed beta-sheet edges, and ligand-binding pockets often present clusters of solvent-exposed nonpolar residues. These patches are the strongest air-water-interface attractors.
-
Dipole moments and asymmetric charge. A protein with a strong net dipole tends to align with the interface's electric double layer. Highly anisotropic charge distributions produce reproducible orientational biases even without hydrophobic adsorption.
-
Shape anisotropy. Elongated or disc-shaped particles align their largest cross-section parallel to the air-water interface during thinning. A flat coin lands face-up.
-
Glycan shielding. Heavily glycosylated proteins (HIV Env, many viral spikes) can be shielded from denaturation but still adopt preferred orientations because glycans cluster on specific faces.
Glaeser & Han (2017) reviewed the biophysics of this process in detail and concluded that the air-water interface should be considered a default failure mode, not an edge case (Glaeser & Han, 2017).
A Brief Catalogue of Common Failure Modes
In our experience surveying difficult cryo-EM projects, the same handful of pathologies recur:
-
The pancake. A flat, oblong protein lies flat on the interface. You see only top views.
-
The lollipop. A protein with a small hydrophobic head pinned to the interface and a long stalk hanging into solution. Strong top-and-bottom bias, with rotational ambiguity around the stalk axis.
-
The dipole. A protein with a marked net dipole moment aligns predictably with the interface field. Two orientations dominate, 180° apart.
-
The denaturer. Particles that fully or partially unfold on contact with the interface. 2D classes look fragmented; the "good" particles are an under-represented subset that vitrified before contact.
-
The shielded mosaic. Glycoproteins where some faces are glycan-protected and others are not. Mixed orientations, hard to interpret.
Each failure mode has different best fixes. The diagnostics below help you decide which one you have.
Diagnostic Tools: How to Tell What You Are Looking At
You can only fix preferred orientation if you correctly characterize it. Global resolution numbers will mislead you. Use these four diagnostics.
Diagnostic 1: Euler Angle Distribution Plots
Every refinement package (RELION, cryoSPARC, CisTEM) produces an angular distribution plot—a sphere or Mollweide projection showing particle counts per Euler bin.
What to look for:
-
Uniform speckle: Healthy. Particles populate orientation space evenly.
-
A bright spot or two: Strong preferred orientation. One or two views dominate.
-
A ring or great circle: Common for elongated particles—rotation about the long axis is sampled but tilt is not.
-
A bald patch: Missing views. The reconstruction has a missing cone.
Counting particles is not enough; one must also weight by the contribution to Fourier space. A million top views contribute zero new information beyond the first ten thousand.
Diagnostic 2: 3D FSC and Directional Resolution
Standard FSC computes a spherically averaged resolution. It is blind to anisotropy. The 3D FSC method (Tan et al., 2017) computes the FSC in cones of directions and reports the full angular dependence of resolution.
A standard FSC curve gives you one number—say, 3.2 Å at FSC = 0.143. The 3D FSC instead samples that same threshold across many directions on the sphere and reports the distribution of crossing points. A protein with severe top-view bias might show 2.8 Å across the equator and 4.5 Å along the polar axis: the polar direction is sampled badly, and the map is correspondingly smeared along Z.
Critically, the 3D FSC sphere visualization shows you which directions are bad, not just how bad they are. This is essential for choosing a fix: a missing cone along Z calls for tilting; a missing equatorial ring suggests something fundamentally different (rare, but real for some elongated specimens).
What to look for:
-
3D FSC sphere: Uniform = isotropic resolution. Dimpled = anisotropic.
-
Sphericity index: A single number (0-1) summarizing how isotropic the resolution is. >0.9 is good, <0.7 is a serious anisotropy problem.
-
Resolution along Z vs XY: If Z resolution is dramatically worse, you have a missing cone in the Z direction (i.e., side views are missing).
Diagnostic 3: Per-View 2D Class Counts
Before 3D refinement, examine the 2D classification populations. If one class contains 80% of particles and looks like a top view, you have a problem. This signal is visible after the first hour of analysis—do not wait for a 3D map to discover it.
Two pathologies hide here. The first is the obvious one: one class dominates. The second is more subtle: 2D classes look diverse, but they are actually rotations of the same view rather than distinct projections. Watch for many classes that share the same overall outline but differ only in in-plane rotation—that is rotation around the optical axis, not new orientation information.
Diagnostic 4: cryoEF and Sphericity
Naydenova & Russo (2017) introduced the "efficiency" metric: the ratio of effective Fourier-space sampling achieved versus uniform sampling (Naydenova & Russo, 2017). Their cryoEF tool quantifies the impact of an angular distribution on reconstruction quality.
Interpretation:
| Efficiency | Interpretation | Action |
|---|---|---|
| > 0.8 | Excellent sampling | Proceed |
| 0.6 - 0.8 | Acceptable, minor anisotropy | Likely fine for most purposes |
| 0.4 - 0.6 | Significant bias | Map will be noticeably anisotropic |
| < 0.4 | Severe bias | Reconstruction will be unreliable |
Diagnostic Signature Table
| Signature | Likely Underlying Cause |
|---|---|
| Single bright Euler spot, intact 2D classes | Hydrophobic face pinned at interface; particle intact |
| Single spot, but 2D classes show fragmented density | Denaturation at interface; partial unfolding |
| Ring in Euler space, elongated 2D | Anisotropic shape; long axis parallel to interface |
| Missing cone along Z, otherwise uniform | Classic "top-view bias" from flat particles |
| Two opposite spots (180° apart) | Dipolar alignment in the interface field |
| Heterogeneous: some uniform, some biased | Multiple conformers with different surface chemistry |
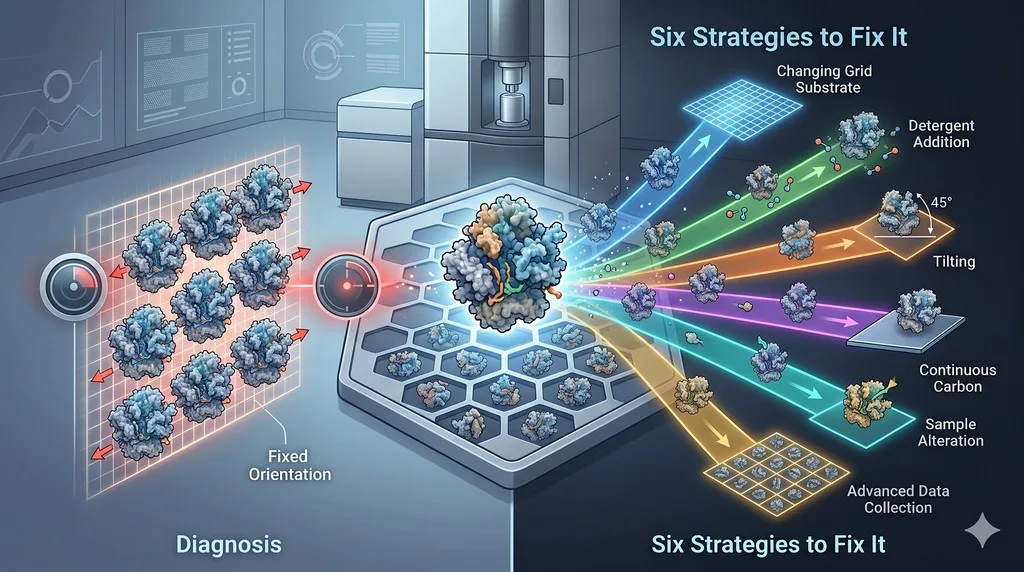
Strategy 1: Tilted Data Collection
The idea: collect data at a stage tilt of 20-40°, mixing additional views into the dataset without changing the sample.
Tan et al. (2017) systematized this approach and showed that tilting can rescue anisotropic reconstructions from previously unusable samples, including hemagglutinin trimer—a notorious top-view protein (Tan et al., 2017).
Pros:
-
No biochemistry. Apply on existing grids in hours.
-
Often the fastest path to a usable map.
-
Compatible with all standard data-collection software.
Cons:
-
Increased ice thickness in the beam (1/cos θ) reduces SNR.
-
Beam-induced motion and charging are worse at high tilt.
-
CTF estimation per particle is harder (defocus gradient across the field).
-
Throughput drops; useful particles per square decline.
-
For severe bias, even 40° tilt may not fully fill in the missing cone.
When to choose: moderate preferred orientation (sphericity 0.6-0.8); time-critical; sample is precious; biochemistry is already optimized.
Practical tips:
-
Collect a small dataset at 30° first, before committing the full session.
-
Reduce dose per frame to compensate for the longer exposure path in tilted ice.
-
Use defocus values toward the lower end (-0.8 to -1.5 µm) to reduce CTF-aliasing artifacts on tilted images.
-
Be ready to mix tilted and untilted data in the final reconstruction—the untilted views still anchor the alignment.
-
Apply per-particle CTF refinement after initial 3D refinement; this can recover 0.3-0.5 Å of resolution on tilted data.
-
Beam-image-shift workflows (using multi-shot with image shift) require careful calibration at high tilt—local beam-tilt corrections matter more than usual.
Strategy 2: Detergent and Surfactant Additives
The idea: add a mild surfactant at low concentration to compete with the protein at the air-water interface, blocking particle adsorption.
The best-studied additives:
-
CHAPSO (zwitterionic, 4-8 mM): widely used; rescued multiple difficult proteins including spliceosomal complexes and pyruvate dehydrogenase (Chen et al., 2019).
-
Fluorinated octyl maltoside (fos-octyl maltoside) and other fluorinated detergents: very low surface tension, often effective when CHAPSO fails.
-
DDM, LMNG: detergents used for membrane proteins can incidentally help with orientation if used carefully.
-
n-octyl-β-D-glucoside (β-OG): classic, but can disrupt some complexes.
Pros:
-
Cheap, fast, no engineering required.
-
Often gives an immediate readout on the next freezing session.
-
Can be combined with other strategies.
Cons:
-
Some additives destabilize complexes or membrane proteins.
-
Trial-and-error: there is no universally predictive theory for which additive works for which protein.
-
Can alter ice thickness and particle distribution unpredictably.
-
May change protein conformation (CHAPSO is mildly chaotropic).
When to choose: hydrophobic-patch-driven orientation; soluble proteins; willing to screen 4-8 conditions; complex tolerates mild surfactants.
A standard screen to start with:
| Additive | Concentration | Notes |
|---|---|---|
| No additive (control) | — | Baseline |
| CHAPSO | 4 mM, 8 mM | Most-tested rescue |
| Fluorinated octyl maltoside | 0.01-0.05% (w/v) | Try when CHAPSO underwhelms |
| β-OG | 0.1% | Older, occasionally helpful |
| DDM | 0.005% | For mildly hydrophobic samples |
| Tween-20 | 0.005% | Last resort, can denature |
Run each on a single grid, image briefly (50-200 movies), and check 2D class distributions before committing to a full collection.
Strategy 3: Surface Mutagenesis
The idea: redesign the protein surface to remove the hydrophobic patches or dipoles that drive interface adsorption.
This is the most rational approach but also the most expensive in time. It requires:
-
Identifying solvent-exposed hydrophobic clusters not involved in function.
-
Designing point mutations (typically Leu/Ile/Val/Phe → Ser/Thr/Lys/Glu).
-
Ensuring mutations do not destabilize the protein (ΔΔG analysis).
-
Re-expressing, purifying, validating activity, and refreezing.
For dipolar bias, one introduces compensating charges on the opposite face, or removes the dominant charge cluster.
Pros:
-
When it works, it works durably—the rescued construct can be used by every group thereafter.
-
Addresses the root cause, not just the symptom.
-
Often improves stability and yield as a side effect.
Cons:
-
2-6 months of work per construct.
-
Risk of compromising activity or fold.
-
Requires expression infrastructure and biophysical characterization.
-
Hard to do without computational support; brute-force screening is wasteful.
When to choose: high-value target you will work on for years; orientation bias is severe and tilting plus detergents have failed; you have or can predict the relevant surface chemistry.
Design heuristics for surface mutations:
-
Target solvent-accessible hydrophobic clusters > 200 Ų that are not at functional interfaces.
-
Prefer Leu/Ile/Val → Ser or Thr (small polar) over the more disruptive → Lys/Glu, which can alter charge.
-
Avoid residues participating in long-range hydrophobic packing (often visible as low B-factor in a homologue).
-
Validate predicted ΔΔG < +1.5 kcal/mol per mutation; combine 2-4 mutations cautiously.
-
Always check that the new surface is not predicted to increase aggregation propensity—you can trade one problem for another.
Strategy 4: Fab and Nanobody Fusion
The idea: bind an antibody fragment (Fab) or nanobody to the target. The asymmetric handle breaks orientation bias, increases particle mass for better alignment, and often supplies a fiducial for symmetry-breaking.
This approach was famously enabling for GPCR cryo-EM and remains the default rescue for many small or symmetric targets.
Pros:
-
Adds 25-50 kDa of mass and breaks symmetry simultaneously.
-
Can convert an unusable 80 kDa target into a routine 110-130 kDa one.
-
Mature methodology with established platforms.
-
Pre-existing nanobody libraries (synthetic or immune) are available.
Cons:
-
Requires generating or sourcing a binder—weeks to months of work.
-
Binder must bind a non-functional face if you want to preserve activity.
-
Adds heterogeneity if binding is sub-stoichiometric.
-
For very small targets (<30 kDa), even with a binder, particles may still be hard to align.
When to choose: small targets, symmetric homo-oligomers, GPCRs, ion channels, transporters, or any case where pursuing the binder is faster than redesigning the surface.
Choosing where the binder lands:
-
Avoid the active site if you need to capture the active state.
-
Prefer epitopes that are conformation-stable (not on flexible loops) for crisp reconstruction.
-
For symmetric homo-oligomers, choose an epitope present in only one copy (e.g., via mutational asymmetry) to break the symmetry usefully.
-
For membrane proteins, the extracellular face is usually the most accessible and gives the most useful asymmetric handle.
Strategy 5: Continuous Carbon, Graphene Oxide, and Affinity Supports
The idea: use a support film that adsorbs particles before they reach the air-water interface, decoupling particle orientation from the interface chemistry.
Common supports:
-
Continuous thin carbon (~3-5 nm): the classic solution; reduces interface contact but adds background noise.
-
Graphene oxide: very thin, low background, mildly hydrophilic; often shifts orientation distributions.
-
Pristine graphene: extremely thin and low-noise, but hydrophobic and tricky to make particles stick.
-
Functionalized supports: streptavidin monolayers, Ni-NTA lipid layers, and affinity grids that capture His-tagged or biotinylated particles in defined orientations.
Drulyte et al. (2018) review the influence of supports and additives on particle behavior on grids (Drulyte et al., 2018).
Pros:
-
Often rescues otherwise hopeless samples.
-
Affinity supports can deliberately enforce a desired orientation.
-
Reusable knowledge: once the support works for a class of proteins, it generalizes.
Cons:
-
Background noise reduces achievable resolution (especially with carbon).
-
Graphene oxide is batch-variable.
-
Affinity supports require tag engineering and careful surface chemistry.
-
Some supports themselves induce a new preferred orientation, just a different one.
When to choose: particles that denature at the air-water interface; small particles where any additional signal would help; cases where a tag for affinity grids is already present.
Strategy 6: Alternative Vitrification Methods
The idea: change how the grid is made so that particles spend less time at the air-water interface before vitrification.
Methods:
-
Jet vitrification and ultrasound spraying (Spotiton, Chameleon by SPT Labtech): spray small droplets onto a self-blotting nanowire grid and plunge in <100 ms (Noble et al., 2018).
-
Self-blotting grids (nanowire grids): eliminate filter-paper blotting and its associated time/contamination.
-
Microfluidic systems: deliver protein directly to the grid in defined volumes and timings.
-
VitroJet, cryoWriter, and other dispense-and-plunge instruments: write small volumes directly and plunge rapidly.
Pros:
-
Directly addresses the root mechanism: less time, less interface damage.
-
Has rescued samples where every additive failed.
-
Compatible with most downstream supports and additives.
Cons:
-
Instrument access is limited; not every facility has one.
-
Sample consumption per grid is similar or lower, but optimization can still be sample-heavy.
-
Newer methods are still maturing; reproducibility is improving but not yet universal.
-
Cannot rescue a sample that is already denatured in solution.
When to choose: particles known to denature at the interface; high-value samples worth instrument access; you have already tried conventional blotting and detergents.
Strategy Comparison Table
| Strategy | Time to Test | Cost | Success Rate (single attempt) | Risk to Sample | Reusable |
|---|---|---|---|---|---|
| Tilted data collection | < 1 day | Microscope time | 40-60% | None | Yes (workflow change) |
| Detergent additives | 1-2 days | < $100 | 30-50% | Low (some) | Per condition |
| Surface mutagenesis | 2-6 months | Lab effort | 60-80% if rationally designed | Activity loss | Yes (new construct) |
| Fab / nanobody fusion | 1-6 months | Moderate to high | 70-90% | Activity occlusion | Yes (reagent) |
| Continuous carbon / GO | 1-2 weeks | Modest | 40-70% | Background noise | Per protein class |
| Alternative vitrification | Instrument-dependent | Instrument access | 50-80% | Low if optimized | Yes (workflow) |
A typical practical decision tree: try tilting first (cheap, fast, informative); add detergent screening in parallel; if these fail and the target is high-value, invest in a Fab or surface redesign.
A Realistic Combined Workflow
Most successful rescues we have seen combine two or more strategies rather than relying on a single fix:
-
Day 1: Diagnose with existing data—Euler distribution, sphericity, 2D-class fractions.
-
Day 2-3: Collect a small tilted dataset (30°). If sphericity rises above 0.85, you may be done.
-
Day 3-5: In parallel, freeze grids with CHAPSO and a fluorinated detergent. Compare 2D classes after a quick screen.
-
Week 2: If sample is denaturing, test graphene-oxide grids or affinity-grid capture using an existing tag.
-
Month 1-3: If none of the above suffices, commit to surface redesign or Fab generation based on in-silico predictions of where the hydrophobic patches live.
-
Throughout: Re-run diagnostics after every change. Sphericity and angular efficiency are your scoreboard.
The single biggest mistake is to skip step 1 and start applying fixes without knowing which failure mode you are facing. A pancake protein and a denaturer require different treatments.
What Surface Chemistry Reveals Before You Freeze
The diagnostic and fix loop above is reactive: you only learn there is a hydrophobic patch problem after a failed grid. Much of the wasted effort in cryo-EM optimization comes from not knowing—before you ever pipette protein onto a grid—where the hydrophobic patches are, how dipolar the molecule is, or which face is glycan-shielded.
Modern surface-property prediction tools can map these features from sequence and structure. Orbion's AstraUNFOLD module computes solvent-accessible hydrophobic patches, charge distributions, and disorder propensity across the structure, flagging surfaces likely to drive air-water-interface adsorption. AstraBIND identifies epitope-accessible regions and predicts where a Fab or nanobody could be raised to provide an asymmetric handle without occluding the functional site. The Construct Design module lets users introduce surface mutations, fusion tags, or affinity-grid handles in silico and evaluate stability impact (via AstraDDG) before committing to wet-lab construction.
The result is that "preferred orientation" stops being a post-hoc diagnosis and starts being a design parameter—one you optimize alongside expression and stability before you book the Krios. The same predictions also inform which Fab epitope to raise, which face to glycan-engineer, and which fusion strategy is least likely to compromise activity.
For groups working on long-term cryo-EM targets, this shift from reactive to proactive surface design is the same kind of transition crystallographers made when they began screening surface entropy reduction mutants instead of brute-force re-screening crystallization conditions. The technique repays the up-front investment many times over for any target that will be re-frozen across multiple states, ligands, or conformations.
The Bottom Line
| Question | Answer |
|---|---|
| Is preferred orientation a resolution problem? | No—it is a sampling problem. Global FSC can be high while the map is uninterpretable. |
| What is the dominant cause? | Adsorption to the air-water interface, driven by hydrophobic patches, dipoles, or shape. |
| How do you diagnose? | Euler angle distribution, 3D FSC, sphericity index, 2D-class populations. |
| What is the fastest fix? | Tilted data collection (often a same-day rescue). |
| What is the most durable fix? | Surface redesign or a Fab fusion—addresses the root cause. |
| What should you do first? | Diagnose. Do not apply fixes blindly. |
A 3 Å map with sphericity 0.5 is worse, for biological interpretation, than a 4 Å map with sphericity 0.95. Preferred orientation is one of those problems where the right diagnosis saves months.
Quick Reference: Decision Cheatsheet
-
Sphericity > 0.85 and 2D classes look balanced → proceed; you do not have a serious orientation problem.
-
Sphericity 0.65-0.85 → tilt your next collection by 20-30° and re-evaluate.
-
Sphericity < 0.65 with intact particles → screen detergents in parallel with tilting.
-
2D classes show fragmented density → particles are denaturing; move to GO/carbon supports or rapid vitrification.
-
Diagnosed dipolar bias → consider surface charge engineering or Fab fusion at the dipole-aligned face.
-
Small (<80 kDa) symmetric target → start the Fab/nanobody pipeline early; do not wait for a failed dataset to begin.
References
-
Tan YZ, Baldwin PR, Davis JH, Williamson JR, Potter CS, Carragher B, Lyumkis D. (2017). Addressing preferred specimen orientation in single-particle cryo-EM through tilting. Nature Methods, 14:793-796. Link
-
Naydenova K, Russo CJ. (2017). Measuring the effects of particle orientation to improve the efficiency of electron cryomicroscopy. Nature Communications, 8:629. Link
-
Noble AJ, Dandey VP, Wei H, Brasch J, Chase J, Acharya P, Tan YZ, Zhang Z, Kim LY, Scapin G, Rapp M, Eng ET, Rice WJ, Cheng A, Negro CJ, Shapiro L, Kwong PD, Jeruzalmi D, des Georges A, Potter CS, Carragher B. (2018). Routine single particle CryoEM sample and grid characterization by tomography. Nature Methods, 15:793-795. Link
-
D'Imprima E, Floris D, Joppe M, Sánchez R, Grininger M, Kühlbrandt W. (2019). Protein denaturation at the air-water interface and how to prevent it. eLife, 8:e42747. Link
-
Glaeser RM, Han BG. (2017). Opinion: hazards faced by macromolecules when confined to thin aqueous films. Biophysics Reports, 3:1-7. Link
-
Drulyte I, Johnson RM, Hesketh EL, Hurdiss DL, Scarff CA, Porav SA, Ranson NA, Muench SP, Thompson RF. (2018). Approaches to altering particle distributions in cryo-electron microscopy sample preparation. Acta Crystallographica D, 74:560-571. Link
-
Chen J, Noble AJ, Kang JY, Darst SA. (2019). Eliminating effects of particle adsorption to the air/water interface in single-particle cryo-electron microscopy: bacterial RNA polymerase and CHAPSO. Journal of Structural Biology: X, 1:100005. Link