Blog
Orbion Team
Tag-Free Purification Strategies: When Affinity Tags Ruin Your Downstream Assay

Your protein is His-tagged. Your binding assay is for metal-dependent activity. You're either purifying clean protein with bad tags, or fighting the chemistry every step. There's a third way. Tag-free purification is older than affinity chromatography, often cheaper, and—for certain downstream applications—the only path to data you can trust.
The pendulum has swung too far toward affinity tags. They are convenient, but convenience is not free: tags distort assays, perturb structures, and occasionally lie to you about the protein you think you have.
Key Takeaways
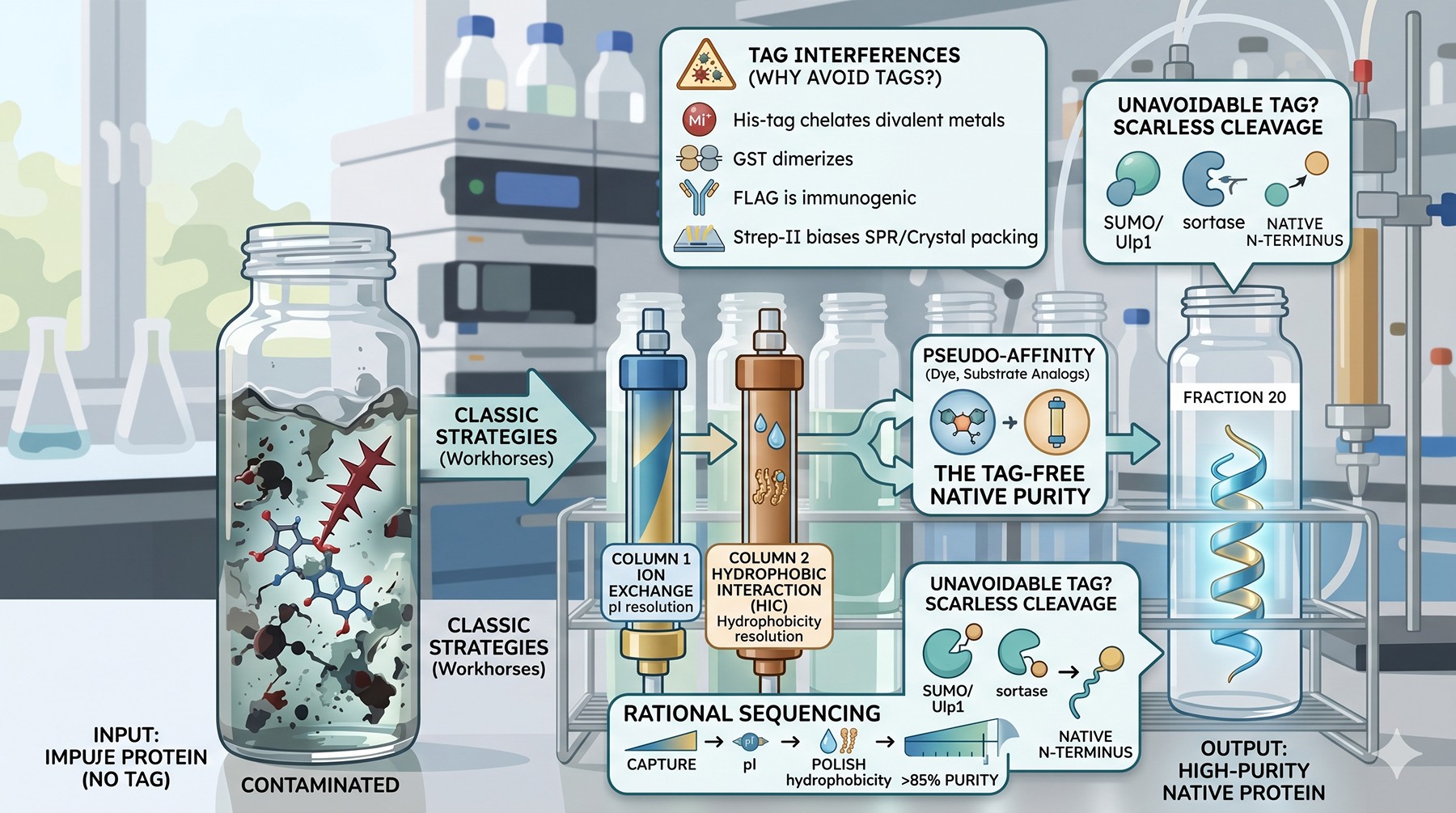
Tags are not inert: His-tags chelate divalent metals, GST dimerizes, FLAG is immunogenic, and even small tags like Strep-II can drift SPR baselines and bias crystal packing
Classic ion exchange and HIC remain the workhorses of tag-free purification—well understood, scalable, and resolving by orthogonal physicochemical properties
Step ordering matters more than step choice: a rational sequence (capture → intermediate → polish) based on pI, hydrophobicity, and MW typically yields >85% purity in three steps
Pseudo-affinity ligands (dye, substrate analog, immobilized metal-without-the-tag) bridge the gap when classical chromatography cannot resolve the contaminant
When some tag is unavoidable, choose chemistries that leave no scar: SUMO/Ulp1, sortase, and self-cleaving inteins deliver native termini that ion exchange or HIC alone cannot
When Tags Fail Your Assay
A tag is a small addition. It is also a covalently attached, charged, sometimes structured polypeptide that the protein did not evolve with. For many assays this does not matter. For some, it ruins everything.
Crystallography Artifacts
Affinity tags introduce conformational heterogeneity at protein termini—exactly where crystal contacts often form. Flexible His-tags rarely crystallize themselves; they tether the structured domain at an unpredictable distance from the lattice and reduce diffraction quality. When they do crystallize, they may drive lattice packing in ways unrelated to native biology (Carson et al., 2007).
The clearer failure mode is structured tags. GST dimerizes (Kd in the low nM range), so a GST-fusion solved by X-ray reveals GST contacts as the dominant crystallographic interaction. MBP at 42 kDa often makes the better crystals than the passenger does—and MBP-mediated lattices are an entirely separate research area, not the structural answer most users want.
The practical rule: for de novo crystallography, either remove the tag completely and verify by mass spectrometry that no residues remain, or pick a passenger-tag pair where the tag participates in productive crystal contacts on purpose (Center for Eukaryotic Structural Genomics-style MBP fusion crystallography is a specialist technique, not a default).
Metal-Dependent Assays
This is the canonical case for tag-free purification. His6 chelates Ni²⁺, Cu²⁺, Zn²⁺, Co²⁺, Mn²⁺—essentially every divalent transition metal a metalloenzyme might need. The consequences:
Activity assays read artifactually low because the tag sequesters the cofactor before it reaches the active site
Apparent Km for metal-substrate complexes shifts by orders of magnitude
Metal stoichiometry by ICP-MS is inflated because His6 brings along its own occupancy
Crystal soaks with metals decorate the tag rather than the active site
For zinc finger proteins, calcium-binding EF-hands, magnesium-dependent kinases, and any metalloenzyme, the His-tag is not a neutral addition—it is a competing ligand. Even after Ni-NTA elution and dialysis, residual Ni²⁺ on the tag can poison subsequent metal-loading experiments (Bornhorst & Falke, 2000).
Immunoassay Interference
Tags are antigens. Two specific failure modes:
Anti-tag antibodies in clinical or pharmacological samples. FLAG, HA, and Myc tags are derived from real epitopes (FLAG from a custom sequence, HA from influenza hemagglutinin, Myc from human c-Myc). Patient sera occasionally contain antibodies that recognize HA or Myc, generating false positives in ELISA or surface plasmon resonance assays.
Cross-reactivity in proteomics. Pulldowns using GST or MBP fusions co-purify endogenous binders of the tag—amylose-related lectins for MBP, glutathione transferases for GST. The contaminant list grows; the real interactor list shrinks under stringent washing.
SPR and BLI Baseline Drift
In surface plasmon resonance, the immobilized analyte must be homogeneous and stably oriented. His-tagged proteins captured on NTA-chips look attractive (oriented, regenerable) but the tag-chip interaction has a measurable koff that drifts the baseline over minutes-to-hours. Kinetic fits become unreliable; equilibrium analysis becomes the only option.
Strep-II tags (8 residues, WSHPQFEK) have similar issues on Strep-Tactin surfaces. For kinetic SPR, covalent amine coupling of tag-free protein is the gold standard, even if it sacrifices orientation control.
In Vivo Immunogenicity
For in vivo use—diagnostic reagents, vaccine antigens, therapeutic candidates—tags are immunogenic liabilities. His6 itself is weakly immunogenic in mice and humans; GST and MBP are highly immunogenic and are sometimes deliberately used as carrier proteins for that reason. For any therapeutic candidate, regulatory pathways effectively require complete removal of expression and purification tags.
A subtler problem: even after "complete" tag removal, residual fusion protein contaminates the final product at low percentages. For research, 1% contamination is invisible on a Coomassie gel. For clinical use, 1% contamination is a regulatory document. Tag-free purification from the start avoids this entire conversation.

NMR and Biophysics
For NMR spectroscopy, tags add resonances that overlap with the target's signals and complicate assignment. A 42 kDa MBP fusion of a 10 kDa target is dominated by MBP signals; the target spectrum is buried under a forest of unwanted peaks. Isotope-labeled MBP can theoretically be filtered out by perdeuteration of the fusion partner, but the additional cost (~$2000 per labeled prep) makes tag-free production attractive even at small scale.
Analytical ultracentrifugation (AUC), differential scanning calorimetry (DSC), and isothermal titration calorimetry (ITC) all benefit from tag-free protein. Tags add mass that biases sedimentation coefficients, add unfolding transitions that complicate DSC, and add binding heat that contaminates ITC baselines.
Classic Tag-Free Strategies
These methods predate affinity chromatography. They remain the foundation of biopharmaceutical-scale purification and are perfectly suitable for research-scale work when applied thoughtfully (Scopes, 1994).
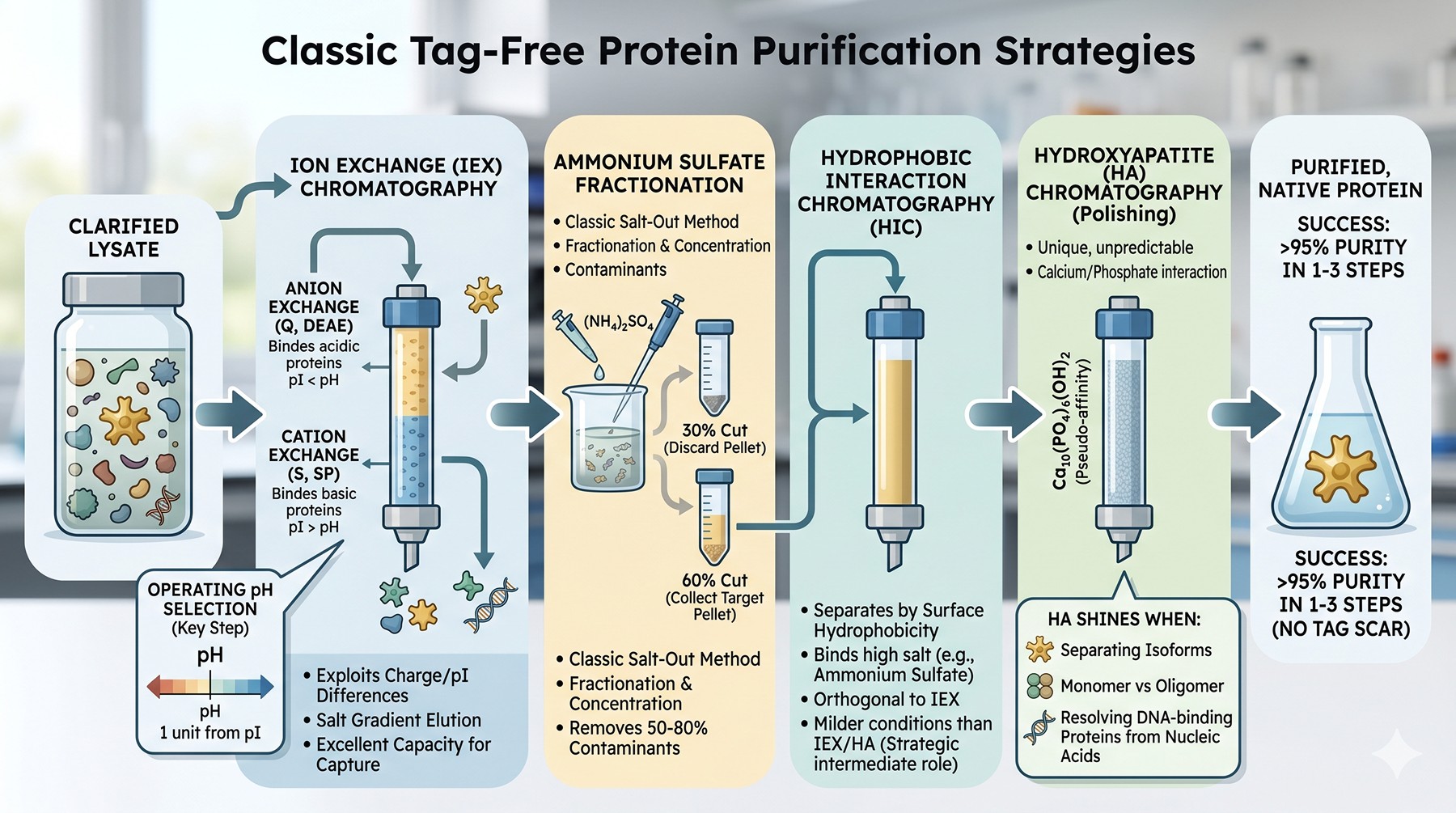
Ion Exchange Chromatography
The most powerful general method for separating proteins. IEX exploits the surface charge differences that arise from differing pI values.
Anion exchange (Q, DEAE, ANX): binds proteins more acidic than the operating pH (pI < pH).
Cation exchange (S, SP, CM): binds proteins more basic than the operating pH (pI > pH).
Operating pH selection is the single most important decision. Set the pH ~1 unit away from your target's pI on the side that gives binding. For a target with pI 5.8, anion exchange at pH 7.5 gives strong binding; cation exchange at pH 4.5 also works but may be too acidic for stability. The chosen pH should be inside the protein's stability window (verified by thermal shift or short stability test).
Resolution from contaminants depends on the pI distribution of host cell proteins. E. coli lysate has a bimodal pI distribution, with peaks near pH 5.5 and pH 9.5. Targets near these peaks will be hard to resolve from the bulk; targets in the valleys (pH 6.5–8.5 or pH 10+) resolve well.
Salt gradient elution typically uses NaCl 0 → 1 M over 10–20 column volumes. Steep gradients give higher yield, shallow gradients give higher resolution. For tag-free polishing, shallow gradients (over 30+ CV) routinely deliver >95% purity in a single step.
Capacity is excellent (50–150 mg protein/mL resin for modern monolithic and bead-based resins), supporting capture from clarified lysate at preparative scale.
Hydrophobic Interaction Chromatography (HIC)
HIC separates by surface hydrophobicity. Protein binds at high salt (typically 1–2 M ammonium sulfate); elution is achieved by lowering salt. Selectivity is orthogonal to ion exchange—two proteins of similar pI can have very different hydrophobicity.
Ligand choice ranges from weakly hydrophobic (ether, butyl) to strongly hydrophobic (octyl, phenyl). Modern resins (Capto Phenyl, Toyopearl Butyl-650) are versatile starting points. The general rule: stronger ligand = lower starting salt needed = milder conditions; but stronger ligand may also fail to release tightly-bound protein.
Strategic role: HIC is often the cleanest intermediate step in a three-step purification. The protein arrives in high-salt buffer directly from ammonium sulfate fractionation or after a salt gradient elution from anion exchange—no buffer exchange required.
Caveats: some proteins denature at high salt or on hydrophobic surfaces. Membrane-associated proteins and proteins with exposed hydrophobic patches may aggregate. Always run a small-scale HIC test before committing.
Hydroxyapatite (HA) Chromatography
Hydroxyapatite (Ca₁₀(PO₄)₆(OH)₂) is a pseudo-affinity matrix that interacts with proteins through both electrostatic and coordination chemistry. Carboxylate groups bind to calcium sites; basic groups bind to phosphate sites. Resolution is unpredictable from first principles—HA simply works differently for every protein.
When HA shines:
Separating closely related isoforms that IEX and HIC cannot resolve
Separating monomer from oligomer (HA is exquisitely sensitive to quaternary structure)
Resolving DNA-binding proteins from contaminating nucleic acids (DNA elutes with phosphate; protein elutes with sodium phosphate at intermediate concentration)
Buffer: typically a phosphate gradient (10–500 mM sodium phosphate, pH 6.5–7.5). Calcium-containing additives must be avoided; chelators (EDTA) destroy the column.
Practical note: HA is fragile, expensive, and slow. Use it as a polishing step, not for capture.
Ammonium Sulfate Fractionation
The oldest scalable separation method, and still useful. Proteins precipitate at distinct salt concentrations based on surface hydrophobicity (Hofmeister salting-out). Two cuts—a "remove" cut (lower percentage, discard precipitate) and a "collect" cut (higher percentage, retain precipitate)—can concentrate target 5–10x while removing 50–80% of contaminating proteins.
Typical workflow: lysate → centrifuge → 30% ammonium sulfate (discard pellet) → 60% ammonium sulfate (collect pellet) → resuspend in HIC equilibration buffer → load HIC.
The output of ammonium sulfate fractionation goes directly onto HIC with no buffer exchange. This pairing is the classical "salt-out, salt-on, salt-off" capture strategy that built mid-century enzymology.
Isoelectric Precipitation
At its pI, a protein has zero net charge and minimum solubility. Slowly titrating lysate to the target's pI causes selective precipitation; centrifugation collects the protein. The cleared supernatant retains contaminants with different pIs.
Limitations: many proteins denature or aggregate irreversibly at their pI. The method requires the target to be unusually stable at low net charge (often acidic milk and storage proteins—the foundational targets of the method).
Modern niche: initial bulk fractionation of crude extracts where partial enrichment is sufficient before a chromatographic step.
Modern Tag-Free Strategies
Pseudo-Affinity: Dye Ligand Chromatography
Reactive textile dyes (Cibacron Blue F3GA, Procion Red HE-3B, Reactive Green 19) bind to dinucleotide-binding folds—the Rossmann fold, kinase ATP sites, and NAD(P)-binding domains. The structural basis is decades-old: Cibacron Blue is a planar polyaromatic with sulfonate groups that mimics ADP/NAD geometry (Stellwagen, 1990).
Practical use:
Capture of dehydrogenases, kinases, polymerases, albumins
Resolution of HSA from biological fluids (HSA binds Cibacron Blue with sub-µM affinity)
Negative selection: removing HSA contamination from antibody preparations
Caveats: the binding mechanism is empirical. Two related enzymes may bind to different dyes; the same enzyme may bind tightly to one dye and not another. Screening (commercial mixed-bed PIKSI kits) is the standard approach.
Immobilized Substrate and Substrate Analogs
For enzymes, the most natural affinity is to the substrate itself. Immobilized substrate analogs that resist turnover (transition state analogs, slow-release substrates, dead-end inhibitors) provide single-step purification with extraordinary selectivity:
Immobilized NAD/NADP for dehydrogenases
Immobilized heparin for serine proteases and DNA-binding proteins
Immobilized lectins (Concanavalin A) for glycoproteins
Immobilized calmodulin for CaM-binding enzymes
Immobilized substrate peptides for proteases and kinases
For well-studied targets, commercial resins exist (NAD-agarose, heparin-Sepharose). For novel targets, custom coupling chemistries (NHS-activated Sepharose, CDI-activated resin) allow you to make your own affinity matrix.
Heparin Affinity
Heparin is so widely useful it deserves its own mention. Heparin is a highly sulfated polyanion that binds:
Most growth factors (FGF, VEGF, HGF)
Serine proteases (thrombin, antithrombin)
Many DNA/RNA-binding proteins (because nucleic acids and heparin share polyanionic character)
Lipoproteins
Heparin-Sepharose is a near-universal "second column" for DNA-binding proteins after ion exchange.
Three-Phase Partitioning (TPP)
Add t-butanol and ammonium sulfate to crude extract: protein partitions into a middle phase between an upper t-butanol-rich layer and a lower aqueous-rich layer. Lipids partition up; nucleic acids and salts partition down. TPP achieves simultaneous concentration, partial purification, and detergent/lipid removal in a single step (Dennison & Lovrien, 1997).
Best for: quick concentration of dilute targets; removal of polyphenols from plant extracts; detergent removal from membrane protein preparations.
Aqueous Two-Phase Systems (ATPS)
Polymer-polymer (PEG/dextran) or polymer-salt (PEG/phosphate) systems form two immiscible aqueous phases. Proteins partition between phases based on a combination of hydrophobicity, surface charge, and molecular weight. Tunable, scalable, and gentle.
Use cases: continuous bioprocessing, large-scale antibody capture (commercial PEG/salt extraction is a recurring topic in biopharmaceutical literature), and bulk fractionation of cell lysates.
Why it's underused in research: the multi-parameter optimization (polymer MW, polymer concentration, salt, pH) is more involved than running a chromatography column, and the literature reports are inconsistent. Most groups skip it.

Strategic Purification Design
A multi-step tag-free purification succeeds or fails on the sequence of steps, not the individual choices. The classical hierarchy is:
Capture: high capacity, fast, tolerates crude feed (ion exchange or ammonium sulfate fractionation)
Intermediate purification: orthogonal selectivity (HIC if capture was IEX; IEX if capture was HIC)
Polishing: highest resolution, removes trace contaminants and aggregates (SEC, second IEX, hydroxyapatite)
Ordering by Physicochemical Property
Pick steps that resolve on different properties. Repeating the same separation principle does not help; orthogonality does.
Step Sequence | Properties Used | Typical Application |
|---|---|---|
Anion exchange → HIC → SEC | Charge → hydrophobicity → size | General-purpose tag-free |
Ammonium sulfate cut → HIC → IEX → SEC | Hydrophobicity → hydrophobicity → charge → size | Difficult lysates, low-abundance targets |
Heparin → Anion exchange → SEC | Polyanion affinity → charge → size | DNA-binding proteins, growth factors |
Hydroxyapatite → SEC | Mixed-mode → size | Closely-related isoforms |
Cation exchange → HIC → SEC | Charge → hydrophobicity → size | Basic proteins (pI > 8) |
Calculating Step Yields
Realistic step yields for well-optimized tag-free purifications:
Anion exchange (capture): 70–90%
HIC (intermediate): 60–85%
SEC (polish): 70–90%
Cumulative three-step yield: ~30–70%
For a 1 L culture producing 50 mg/L soluble target, expect 15–35 mg final purified protein. This is lower than a single-step Ni-NTA purification (typical 70–80% yield), and that is the real cost of going tag-free.
Track yields per step, not just final yield. If anion exchange recovers only 30%, the target is not binding (wrong pH, too much salt, or the binding capacity is exceeded by contaminating proteins). If HIC recovers only 30%, the target may be denaturing on the column.

Predicting Contaminant Overlap
Before running the purification, look at the host cell proteome. E. coli has ~4,000 proteins; most fall in the 20–80 kDa, pI 4.5–9 range. Plotting your target on a 2D map (pI vs MW, or pI vs hydrophobicity) identifies whether it sits in a sparse region (easy purification) or a crowded one (difficult).
Tools like Pellegrini-Calace's lab tools (ProtParam, ProtScale) calculate pI and GRAVY (hydrophobicity index) from sequence. For more sophisticated host-proteome overlap analysis, dedicated platforms can map a target's coordinates against the E. coli, Sf9, or HEK293 background proteome.
Working Backwards from Purity Requirements
Different downstream applications have different purity bars. Designing the purification to the bar—rather than to an arbitrary >99%—saves time and protein.
Application | Required Purity | Required Homogeneity | Typical Number of Steps |
|---|---|---|---|
Pull-down / functional screen | >70% | Monomeric or known oligomer | 1–2 |
Antibody production (immunogen) | >85% | Any | 2 |
Enzyme kinetics | >90% | Monomer (or defined state) | 2–3 |
SPR / BLI binding kinetics | >95% | Monomer, no aggregate | 3 (with SEC last) |
Crystallization | >98% | Monodisperse | 3 + buffer optimization |
Cryo-EM | >95% | Defined oligomer | 3 + ice optimization |
Clinical / therapeutic | >99.5% | Strictly defined | 5+ with viral inactivation |
The key insight is that SEC as a polishing step is non-negotiable for any application sensitive to aggregation (SPR, ITC, structural biology). It removes aggregates and confirms monodispersity in a single run.
The Three-Step Heuristic
The "three-column rule" in protein purification: any well-behaved soluble protein can be purified to >95% homogeneity in three chromatographic steps if those steps are chosen for orthogonal selectivity. Cases requiring four or more steps usually have one of:
A persistent co-purifying contaminant (often a chaperone like GroEL, DnaK)
An oligomeric heterogeneity (monomer + dimer + aggregate)
A truncated or proteolyzed variant of the target itself
The first two are addressed by careful step selection; the third requires going back to expression or construct design.
Tag-Free Method Comparison
Method | Resolution | Capacity | Cost | Throughput | Best Role |
|---|---|---|---|---|---|
Anion exchange (Q, DEAE) | High | Very high (50–150 mg/mL) | Low | High | Capture and polish |
Cation exchange (S, SP) | High | Very high | Low | High | Capture, basic targets |
HIC (butyl, phenyl) | Moderate–high | High (20–80 mg/mL) | Low | High | Intermediate step |
Hydroxyapatite | Moderate | Low (5–10 mg/mL) | High | Low | Specialty polish |
Size exclusion (SEC) | Low (1.5x MW resolution) | Very low (1–5% column volume) | Moderate | Low | Polish, aggregate removal |
Ammonium sulfate cut | Low | Unlimited | Very low | High | Pre-chromatography |
Dye-ligand | Moderate (variable) | Moderate | Moderate | Moderate | Pseudo-affinity capture |
Heparin affinity | High (for targets) | Moderate (10–20 mg/mL) | Moderate | Moderate | Specialty capture |
Hydroxyapatite | Moderate | Low | High | Low | Isoform separation |

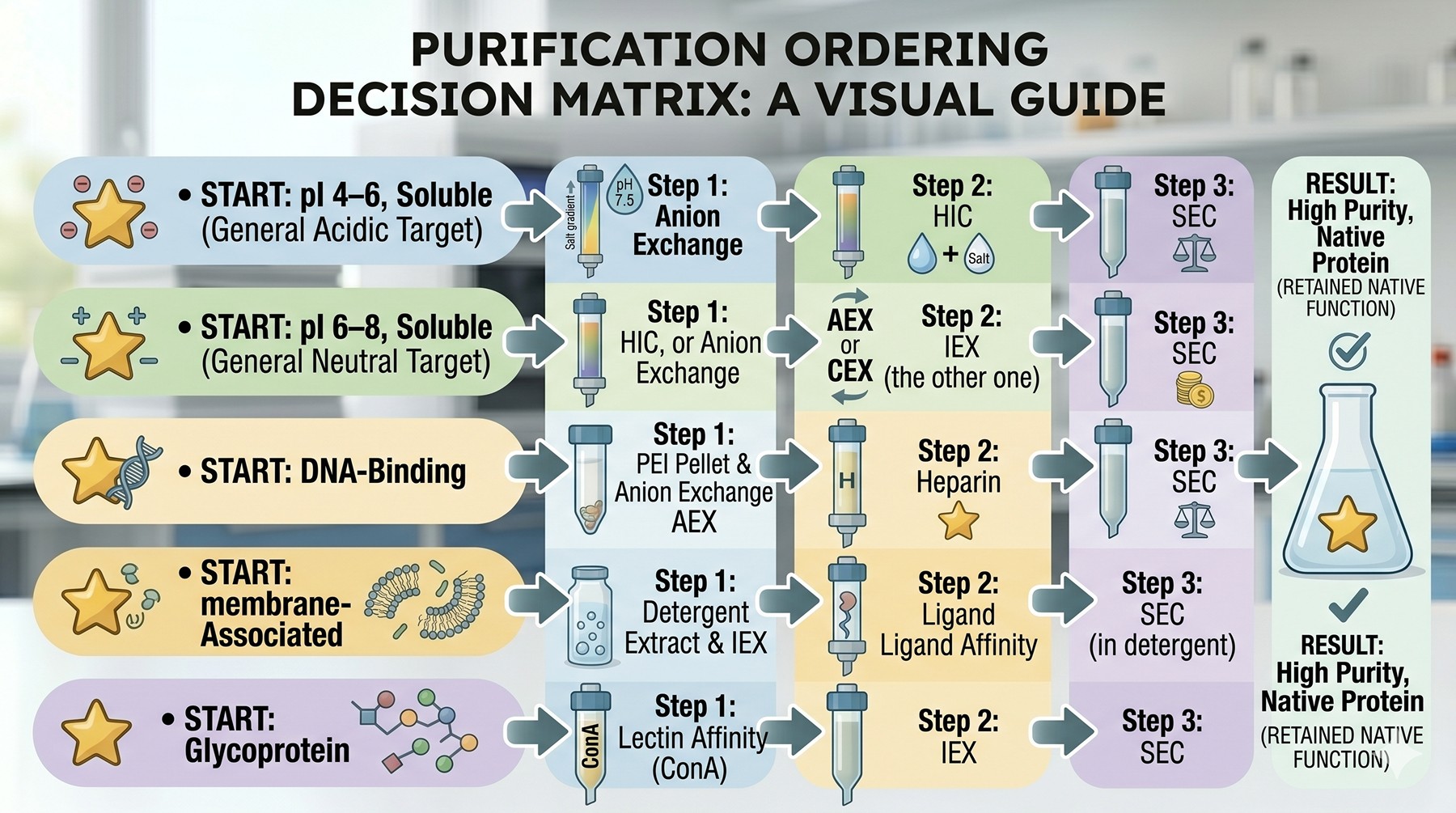
Purification Ordering Decision Matrix
Target Property | Recommended Step 1 (Capture) | Recommended Step 2 | Recommended Step 3 |
|---|---|---|---|
pI 4–6, soluble | Anion exchange at pH 7.5 | HIC | SEC |
pI 6–8, soluble | HIC, or weak anion exchange at pH 8.5 | IEX (the other one) | SEC |
pI 8–10, soluble | Cation exchange at pH 6.0 | HIC | SEC |
pI > 10, basic | Cation exchange | Heparin | SEC |
DNA-binding | Polyethyleneimine pellet then anion exchange | Heparin | SEC |
Has Rossmann fold | Cibacron Blue (dye-ligand) | Anion exchange | SEC |
Membrane-associated | Detergent extraction → IEX in detergent | Ligand affinity if available | SEC in detergent |
Glycoprotein (eukaryotic) | Lectin affinity (ConA) | IEX | SEC |
Closely-related isoforms | Anion exchange shallow gradient | Hydroxyapatite | SEC |
Highly hydrophobic | Ammonium sulfate cut | HIC weak ligand | SEC + glycerol |
When Some Tag Is Unavoidable: Scarless Removal
Sometimes you need the orientation control or yield boost of an affinity tag during capture, but you also need a native protein for your final assay. Modern cleavage chemistries achieve both.
SUMO/Ulp1
SUMO protease (Ulp1) recognizes the tertiary fold of SUMO, not a linear sequence. Cleavage occurs after the C-terminal Gly-Gly of SUMO, generating a completely native N-terminus on the passenger—no extra residues. The reaction is fast (typically 1–2 hours at room temperature), efficient (>95% complete), and exquisitely specific (no off-target cleavage observed).
The catch: SUMO fusion proteins cannot be expressed in eukaryotic systems, because host SUMO proteases (SENPs) will cleave the construct intracellularly before the protein can be harvested. SUMO is a bacterial expression solution.
Sortase A
S. aureus sortase A catalyzes a transpeptidation reaction at an LPXTG motif, cleaving between threonine and glycine and ligating the LPXT to a polyglycine nucleophile. Used for tag removal: the construct is target-LPXTG-affinity_tag; sortase + GG-containing nucleophile cleaves and releases the affinity tag. The target retains an LPXT C-terminal scar (4 residues), or with reverse-orientation constructs, can be released without any C-terminal scar.
Sortase is more commonly used for ligation (attaching labels, fluorophores, lipids) than for tag removal, but the chemistry works in both directions (Mao et al., 2004).
Self-Cleaving Inteins
Inteins are protein domains that catalyze their own excision from a host protein, ligating the flanking sequences with a peptide bond. Engineered "self-cleaving" inteins (Mxe GyrA, Ssp DnaB, Mtu RecA) cleave from one side only, releasing the target protein with a native C-terminus or N-terminus depending on construct orientation.
The IMPACT system (NEB) is the most common commercial implementation: chitin binding domain (CBD) for affinity → intein → target. The construct is captured on chitin beads; cleavage is triggered by adding thiol (DTT or β-ME) or by pH shift. Cleaved native target elutes; CBD-intein remains on the resin (Tan & Wang, 2005).
Caveats: intein cleavage is rarely 100% efficient (typical 60–90%); the conditions for cleavage (pH 6, room temperature, overnight) may not suit unstable targets; premature cleavage during expression reduces yield.
TEV and 3C: Workhorses Despite Scars
For all the elegance of SUMO, sortase, and inteins, the practical reality is that most labs run TEV protease as their default cleavage step. TEV is cheap (express your own), works at 4°C (compatible with unstable targets), and is highly specific (ENLYFQ↓S/G recognition, >99% cleavage at the canonical site, off-target cleavage rare).
The cost is a single Ser or Gly residue left at the N-terminus of the target. For 90% of biochemical applications, this scar is invisible. For crystallography, it adds one residue that may or may not order in the crystal; usually it does not, and is invisible in the electron density.
HRV 3C protease (also called PreScission protease in its GST-fusion commercial form) has a similar profile, leaving Gly-Pro as a 2-residue scar. 3C works better at 4°C than TEV does, making it the preferred choice for cold-sensitive targets.
On-column cleavage is a useful trick when working with TEV or 3C: load the fusion onto the affinity column, wash, then incubate with protease in column buffer. The cleaved target elutes; the affinity tag remains bound. This avoids the reverse-affinity step and saves time.
Cleavage Decision Tree
Requirement | Recommended Strategy |
|---|---|
Native N-terminus, fast cleavage, bacterial expression | His6-SUMO + Ulp1 |
Native N-terminus, eukaryotic expression | TEV (1 residue scar) or HRV 3C |
Native C-terminus | Mxe intein with thiol-induced cleavage |
No scar at either terminus | Tag-free purification from the start |
Tagged for one step only, then released | Intein-CBD (IMPACT) or sortase |

When to Skip Tags Entirely
Despite all the cleavable tag chemistries, sometimes the best answer is to design a tag-free construct from the outset. Indicators:
The target is naturally abundant (>1% of total host protein expression). Tags are unnecessary; classical chromatography is faster.
The target has an intrinsic property exploitable for purification: thermostability (heat-shock the lysate), pI extremes (single IEX captures), specific ligand binding (immobilize the ligand).
The target is for in vivo use (vaccine antigen, biologic precursor). Tag-free from the start avoids the regulatory and immunogenicity questions about residual tags.
The target is being produced at industrial scale (>1 g per batch). Resin costs and tag-removal protease costs become prohibitive; classical chromatography is the only scalable option.
You have time. A tag-free purification takes longer to develop (3–6 weeks of method development) but runs reliably for years thereafter; an Ni-NTA capture takes a day to develop but creates ongoing tag-removal and assay-interference problems.
Common Tag-Free Pitfalls
Pitfall 1: Loading Crude Lysate Onto SEC
SEC is a polishing step, not a capture step. It has tiny capacity (typical loading volume <5% of column volume), is slow, and will be fouled by lipids and nucleic acids from crude lysate. Always do a chromatographic capture (IEX, HIC, or affinity) before SEC.
Pitfall 2: Choosing the Wrong pH for IEX
Sequence-predicted pI is an approximation. Modeled pI calculators (ProtParam, EMBOSS pepstats) assume each ionizable group is independent and uses standard pKa values—both assumptions break down for proteins with buried or networked ionizable residues. Empirical pH-binding tests (load aliquots at pH 5, 6, 7, 8; check flow-through fraction for the target) take a day and prevent weeks of fruitless gradient development.
Pitfall 3: Adding EDTA During Hydroxyapatite
Hydroxyapatite is calcium-based. Chelators destroy it. Even trace EDTA carried over from previous purification steps (a common lysis buffer additive) can ruin an expensive HA column. Always exchange into phosphate buffer before loading HA.
Pitfall 4: Underestimating Ammonium Sulfate Cleavage
Storing protein in 60% ammonium sulfate is standard practice (high stability, near-indefinite shelf life). But ammonium sulfate stocks contain trace metal ions; over weeks at 4°C, redox-active metals can catalyze oxidation of cysteines. For long-term storage of cysteine-containing proteins, switch to a glycerol-based buffer at -80°C.
Pitfall 5: Conflating Step Yield with Step Performance
A 95% step yield does not mean a successful step. If the contaminant load also passed through at 95%, you have not purified anything—you have only concentrated the mixture. Always evaluate steps by purity gain, not just by recovery. SDS-PAGE plus a relative-purity metric (densitometry or label-free MS) tracks this properly.
Pitfall 6: Ignoring Aggregates Until the Final SEC
Aggregates that form during expression and capture persist through all subsequent steps unless explicitly removed. Most ion exchange and HIC columns do not resolve monomeric target from soluble aggregate (the surface chemistry is dominated by the same residues). Only SEC, or a very shallow IEX gradient with native-mass detection, separates aggregate from monomer. Plan for SEC; do not hope your final IEX run also removed aggregates.

The Bottom Line
Situation | Recommended Strategy |
|---|---|
Metalloenzyme, metal-dependent assay | Tag-free from start: IEX → HIC → SEC |
Crystallography of small target | Tag-free, or removable via Ulp1/intein |
SPR kinetics required | Tag-free, amine-couple to chip |
In vivo or therapeutic use | Tag-free, validate by intact-mass MS |
High-yield therapeutic enzyme | Tag-free with substrate-affinity capture |
Rapid expression screening | Affinity tag (His6, Strep), accept the limitations |
Pull-down / interactome | GST or biotinylation tag, not tag-free |
Tag-free unavoidable, low yield | His6-SUMO + Ulp1 + reverse Ni-NTA |
Want native N- and C-termini | Sortase + intein combination, or engineer with tagless construct |
The tag-or-no-tag decision is not a default. It is a function of your downstream assay, your scale, your timeline, and the chemistry of your protein. The wrong choice silently degrades data quality for the entire project lifetime.

Rational Tag-Free Strategy with Orbion
Designing a tag-free workflow requires upfront prediction of the target's physicochemical properties and likely contaminant overlap—exactly what AI-driven characterization is for. Orbion's AstraBIND identifies predicted binding partners and ligand interaction sites, which directly informs pseudo-affinity capture strategy: if your target has a confidently predicted nucleotide-binding fold, Cibacron Blue is on the table; if it has a heparin-binding signature, heparin-Sepharose is a strong first column.
AstraSUIT scores expression-system suitability, flagging whether classical bacterial purification is even appropriate or whether eukaryotic expression (and the corresponding host-proteome overlap) is needed. The Bench module generates step-by-step purification protocols from these predictions—including buffer compositions for IEX (pH choice based on predicted pI), HIC (salt concentration based on predicted surface hydrophobicity), and SEC (column choice based on predicted MW and oligomeric state). Combined with predicted contaminant overlap against the E. coli or HEK293 proteome, this lets you order columns rationally before any wet-lab work begins—turning a 6-week method-development cycle into a 1-week protocol validation.
References
Carson M, Johnson DH, McDonald H, Brouillette C, DeLucas LJ. (2007). His-tag impact on structure. Acta Crystallographica D, 63:295-301. PMC2330224
Bornhorst JA & Falke JJ. (2000). Purification of proteins using polyhistidine affinity tags. Methods in Enzymology, 326:245-254. PMC2774729
Scopes RK. (1994). Protein Purification: Principles and Practice. 3rd ed. Springer-Verlag, New York. Link
Burgess RR. (2009). Refolding solubilized inclusion body proteins. Methods in Enzymology, 463:259-282. DOI
Stellwagen E. (1990). Chromatography on immobilized reactive dyes. Methods in Enzymology, 182:343-357. PubMed
Dennison C & Lovrien R. (1997). Three phase partitioning: concentration and purification of proteins. Protein Expression and Purification, 11(2):149-161. PubMed
Tan LP & Wang R. (2005). Recombinant protein purification using self-cleaving intein tags. Journal of Chromatography A, 1073(1-2):3-15. PubMed
Mao H, Hart SA, Schink A, Pollok BA. (2004). Sortase-mediated protein ligation: a new method for protein engineering. Journal of the American Chemical Society, 126(9):2670-2671. DOI
Book a 20-Minute Demo
Sign up free for unlimited Overview runs — summary, sequence-based analysis, homology search. For the full Characterization — PTMs, binding sites, stability variants, construct design — book a demo and we'll run your target live.
